*2.1. Bacterial Strains and Growth Conditions*
*E. coli* K12 DH5α and JM101(DE3) cells were used for DNA manipulation and expression of carotenoid biosynthesis genes, respectively. These *E. coli* strains and their transformants were grown in 2 × YT (2YT) medium (16 g/L of tryptone, 10 g/L of yeast extract, 5 g/L of NaCl) containing 10 mg/L of tetracycline (as needed) at 37 °C or 21 °C.
*2.2. DNA Isolation of Planococcus maritimus Strain iso-3*
Genomic DNA was prepared from *P. maritimus* strain iso-3 according to the method described by Nishida et al. [
10].
*2.3. Cloning of the crtP (cruO) and aldH Genes from P. maritimus Strain iso-3*
Based on the sequences of the *crtP* and *aldH* genes in the genomes of *Planococcus faecalis* and *Planococcus plakortidis*, primers containing the restriction sites were designed, as shown in Table S1. The coding regions of individual genes were amplified by PCR (polymerase chain reaction) using genomic DNAs. PCR products were cloned into a plasmid vector and sequenced as described [
3].
*2.4. Sequence Analysis*
Homology search was performed using BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Amino acid alignment was performed using MAFFT(http://www.mafft.ccbrc.jp/) and phylogenetic tree was constructed using the neighbor joining method, both as implemented on the GenomeNet (https://www.genome.jp/tools/ete/).
*2.5. Expression of the crtP (cruO) and aldH Genes from P. maritimus Strain iso-3 in E. coli*
The coding region of the *crtP* (*cruO*) gene from *P. maritimus* strain iso-3 was inserted into the plasmids pAC-HIMNFNb and pAC-HIMNFNbG [
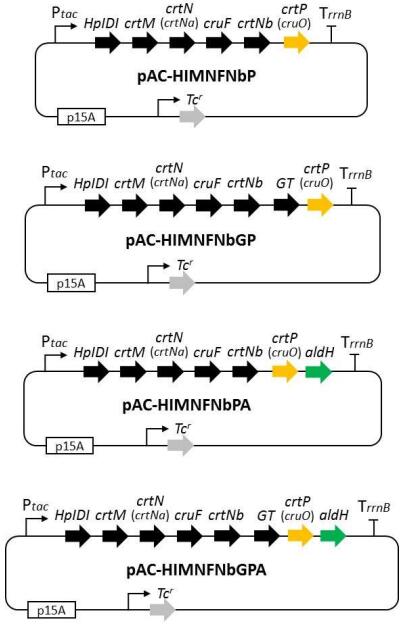
3]. These plasmids were designated pAC-HIMNFNbP and pAC-HIMNFNbGP, respectively (). The coding region of the *aldH* gene from *P. maritimus* strain iso-3 was independently inserted into pAC-HIMNFNbP and pAC-HIMNFNbGP. The resultant plasmids were named pAC-HIMNFNbPA and pAC-HIMNFNbGPA (). All plasmids were introduced into wild-type *E. coli* (JM101 (DE3)). The transformed *E. coli* strains were grown in 2YT medium at 37 °C. The following day, the culture was inoculated in a new 2YT medium (100 ml medium in a 500 ml Sakaguchi flask) and cultured at 21 °C for two days.
. Structure of plasmids constructed in this study.
*2.6. Extraction and High-Performance Liquid Chromatography (HPLC) Analysis of Carotenoids from E. coli Cells*
Extraction of carotenoids from recombinant *E. coli* was performed as described by Fraser et al. [
11]. *E. coli* cultures were centrifuged (8000*g* × 5 min). Cell pellets were extracted with methanol (MeOH) using a mixer for 5 min. Tris–HCl (50 mM, pH 7.5) and 1 M NaCl were added and mixed. Chloroform was added to the mixture and mixed for 5 min. After centrifugation, the chloroform phase was collected and dried through centrifugal evaporation. Dried residues were resuspended in ethyl acetate (EtOAc) and subjected to HPLC with a Waters Alliance 2695-2996 (PDA) system (Waters, Milford, MA, USA). HPLC was performed according to the method described by Yokoyama and Miki [
12] using TSKgel ODS-80Ts (4.6 × 150 mm, 5 µm; Tosoh, Tokyo, Japan). Briefly, the extract was eluted at a flow rate of 1.0 mL/min at 25 °C with solvent A [water (H
2O)-MeOH, 5:95] for 5 min, followed by a linear gradient from solvent A to solvent B (tetrahydrofuran-MeOH, 3:7) for 5 min and solvent B alone for 8 min. The produced carotenoids were identified by comparing their retention times and absorption spectra with those of our authentic standards. When the produced carotenoids were not compounds in our authentic standards, as described in the following sections, we isolated the produced carotenoids and determined their structures using high-resolution electrospray ionization mass spectrometry (HRESI-MS) and nuclear magnetic resonance (NMR) analyses.
*2.7. Isolation of Respective Highly Modified C
30-Carotenoids*
2.7.1. 5‑Hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al (
1)
The transformed *E. coli* cells carrying pAC-HIMNFNbP were collected using centrifugation from 2 L culture and extracted with 540 mL acetone-MeOH (7:2) and 600 mL dichloromethane (CH
2Cl
2)-MeOH (1:1) with sonication in a stepwise manner. The combined extract (1140 mL) was concentrated to a small volume *in vacuo* and partitioned with EtOAc/H
2O (300 mL/150 mL). The EtOAc layer containing the produced carotenoids was evaporated to dryness to obtain yellow oil (125.0 mg). *n*-Hexane (3 mL) was added to the oil and sonicated for 3 min to remove fats and fatty acids (×3). The precipitate (10.1 mg) was subjected to preparative ODS HPLC [column: Develosil C30-UG (20 mm × 250 mm, Nomura Chemical, Co. Ltd, Aichi, Japan), solvent: acetonitrile (CH
3CN)-tetrahydrofuran (THF) (3:2), flow rate: 8.0 mL/min, detection: PDA (250–700 nm)]. The peak at *t
R* 9.6 min was collected and concentrated to dryness to afford pure 5‑hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al (
1) (1.1 mg).
2.7.2. 5‑Hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-oic Acid (
2)
Transformed *E. coli* cells carrying pAC-HIMNFNbPA were collected by centrifugation from 2 L of culture and extracted with acetone-MeOH (7:2) and CH
2Cl
2-MeOH (1:1) by stepwise sonication. The extract (1140 mL) was concentrated to a small volume *in vacuo* and partitioned with EtOAc/H
2O (300 mL/150 mL). The EtOAc layer was evaporated to dryness to obtain a yellow oil (113.6 mg). *n*-Hexane (3 mL) was added to the oil and sonicated for 3 min to remove fats and fatty acids (×3). The precipitate (17.9 mg) was subjected to preparative ODS HPLC [column: Develosil C30-UG (20 mm × 250 mm), solvent: 90% (*v*/*v*) CH3CN containing 20 mM H3PO4-THF (3:2), flow rate: 8.0 mL/min, detection: PDA (250–700 nm)]. The peak at *t
R* 9.8 min was collected and concentrated to 5 mL and partitioned between EtOAc/H
2O (100 mL each). The EtOAc layer was concentrated to dryness to obtain pure 5-hydroxy-5,6-dihydro-4,4′-diapolycopen-4′-oic acid (
2) (1.8 mg).
2.7.3. 5‑Glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al (
3)
The transformed *E. coli* cells carrying pAC-HIMNFNbGP were collected using centrifugation from 2 L of culture and extracted with 540 mL acetone-MeOH (7:2) and 600 mL CH
2Cl
2-MeOH (1:1) with sonication in a stepwise manner. The combined extract (1140 mL) was concentrated to a small volume *in vacuo* and partitioned with EtOAc/H
2O (300 mL/150 mL). The EtOAc layer containing the produced carotenoids was evaporated to dryness to obtain a yellow oil (301.7 mg). *n*-Hexane (3 mL) was added to the oil and sonicated for 3 min to remove fatty acids (×3). The precipitate (142.2 mg) was subjected to preparative ODS HPLC [column: Develosil C30-UG (20 mm × 250 mm, solvent: CH
3CN-THF (3:2), flow rate: 8.0 mL/min, detection: PDA (250–700 nm)]. The peak at *t
R* 8.2 min was collected and concentrated to dryness to afford pure 5‑glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al (
3) (1.2 mg).
2.7.4. 5‑Glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-oic Acid (
4)
Transformed *E. coli* cells carrying pAC-HIMNFNbGPA were collected using centrifugation from 2 L of culture and extracted with acetone-MeOH (7:2) and CH
2Cl
2-MeOH (1:1) using stepwise sonication. The extract (1140 mL) was concentrated to a small volume *in vacuo* and partitioned with EtOAc/H
2O (300 mL/150 mL). The ethyl acetate (EtOAc) layer was evaporated to dryness (203.2 mg). n-Hexane (3 mL) was added to the oil and sonicated for 3 min to remove fats and fatty acids (×3). The precipitate (58.9 mg) was subjected to preparative ODS HPLC [column: Develosil C30-UG (20 mm × 250 mm), solvent: 90% (*v*/*v*) CH
3CN containing 20 mM H3PO4-THF (3:2), flow rate: 8.0 mL/min, detection: PDA (250–700 nm)]. The peak at *t
R* 11.2 min was collected and concentrated to 5 mL and partitioned between EtOAc/H
2O (each 100 mL). The EtOAc layer was concentrated to dryness to obtain pure 5-glucosyl-5,6-dihydro-4,4′-diapolycopen-4′-oic acid (
4) (3.2 mg).
*2.8. Singlet Oxygen-Quenching Activity*
For the measurement of singlet oxygen-quenching activity, 80 µL of 25 µM methylene blue and 100 µL of 0.24 M linoleic acid, with or without 40 µL of carotenoid (final concentration, 1–25 µM; each dissolved in ethanol), were added to 5 mL glass test tubes. The tubes were mixed well and illuminated at 7000 lx and 22 °C for 3 h in a Styrofoam box. Subsequently, 120 µL of the reaction mixture was removed and diluted to 3.48 mL with ethanol. The OD235 was measured to estimate the formation of conjugated dienes. OD235 in the absence of carotenoids was measured as a negative control [no singlet oxygen (1O
2)-quenching activity]. The 1O
2-quenching activity of carotenoids was calculated from OD235 in the presence of carotenoids relative to this reference value.
The activity is indicated as the IC50 value, which represents the concentration at which 50% inhibition occurs.
*2.9. Physico-Chemical Properties of Respective Highly Modified C
30-Carotenoids*
2.9.1. 5‑Hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al (
1)
HR-ESI-MS (+) *m*/*z* 455.29275 (C
30H
40O
2Na; cald. for 455.29260, ∆ 0.33 ppm). UV-Vis λmax (ε) in MeOH 254 (9200), 315 (6800), 407 (43000), 428 (62000), 452 (56000).
1H NMR (CDCl
3) δ: 1.14 (3H, s, H-4), 1.14 (3H, s, H-18), 1.90 (3H, s, H-18′), 1.94 (3H, s, H-19), 1.98 (3H, s, H-20), 1.98 (3H, s, H-20’), 2.02 (3H, s, H-19′), 2.32 (2H, d, *J* = 7.6 Hz, H-6), 5.77 (1H, dd, *J* = 7.6, 15.1 Hz, H-7), 6.14 (1H, d, *J* = 12.0 Hz, H-10), 6.22 (1H, d, *J* = 15.1 Hz, H-8), 6.26 (1H, d, *J* = 14.5 Hz, H-14), 6.33 (1H, d, *J* =15.0 Hz, H-14′), 6.36 (1H, d, *J* =14.0 Hz, H-12), 6.43 (1H, d, *J* =12.3 Hz, H-10’), 6.50 (1H, d, *J* =15.0 Hz, H-12′), 6.63 (1H, dd, *J* =12.0, 14.0 Hz, H-11), 6.65 (1H, dd, *J* =10.7, 14.8 Hz, H-7′), 6.65 (1H, dd, *J* =12.5, 15.0 Hz, H-11′), 6.65 (2H, H-15 and H-15′), 6.75 (1H, d, *J* =14.8 Hz, H-8′), 6.94 (1H, d, *J* =10.7 Hz, H-6′), 9.45 (1H, s, H-4′) (Figure S1).
13C NMR (CDCl3) δ: 12.8 (C-19), 12.8 (C-20), 12.8 (C-18′), 12.8 (C-20′), 13.0 (C-19′), 29.2 (C-4), 29.3 (C-18), 47.4 (C-6), 71.0 (C-5), 122.4 (C-7′), 124.4 (C-11′), 124.8 (C-11), 125.3 (C-7), 129.8 (C-15′)
c, 131.0 (C-10), 131.2 (C-15)c, 132.5 (C-14)
b, 134.8 (C-14′)
b, 134.9 (C-9′), 135.4 (C-13)
a, 136.1 (C-13′)
a, 136.6 (C-5′), 137.3 (C-9), 137.6 (C-12), 137.8 (C-10′), 138.7 (C-8), 141.0 (C-12′), 146.1 (C-8′), 149.4 (C-6′), 194.6 (C-4′) (Figure S2).
a,
b,
c: Interchangeable.
2.9.2. 5‑Hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-oic Acid (
2)
HR-ESI-MS (+) *m*/*z* 471.28902 (C
30H
40O
3Na; cald. for 471.28751, ∆ 1.51 ppm). UV-Vis λmax (ε) in MeOH 254 (9200), 315 (6800), 407 (43000), 428 (62000), 452 (56000). 1H NMR (DMSO-*d
6*) δ: 1.06 (3H, s, H-4), 1.06 (3H, s, H-18), 1.87 (3H, s, H-19), 1.88 (3H, s, H-18′), 1.93 (3H, s, H-20), 1.94 (3H, s, H-20′), 1.96 (3H, s, H-19′), 2.18 (2H, d, *J* =7.1 Hz, H-6), 5.77 (1H, td, *J* =7.1, 16.0 Hz, H-7), 6.12 (1H, d, *J* =16.0 Hz, H-8), 6.13 (1H, d, *J* =10.2 Hz, H-10), 6.32 (1H, d, *J* =10.4 Hz, H-14), 6.36 (1H, d, *J* =15.0 Hz, H-12), 6.38 (1H, d, *J* =11.0 Hz, H-14′), 6.53 (1H, dd, *J* =11.2, 15.3 Hz, H-7′), 6.40 (1H, d, *J* =11.3 Hz, H-10′), 6.48 (1H, d, *J* =14.8 Hz, H-12′), 6.66 (1H, dd, *J* =10.2, 15.0 Hz, H-11), 6.68 (1H, dd, *J* =11.3, 14.8 Hz, H-11′), 6.72 (1H, dd, *J* =11.0, 15.0 Hz, H-15′), 6.73 (1H, dd, *J* =10.4, 15.0 Hz, H-15), 7.12 (1H, d, *J* =11.2 Hz, H-6′) (Figure S3).
13C NMR (DMSO-*d
6*) δ: 12.7 (C-20′), 12.8 (C-20), 12.8 (C-19′), 13.0 (C-19), 13.3 (C-18′), 29.4 (C-18), 29.4 (C-4), 47.6 (C-6), 69.5 (C-5), 124.1 (C-7′), 125.2 (C-11), 125.6 (C-11′), 127.2 (C-7), 128.6 (C-5′), 130.3 (C-10), 130.3 (C-15′), 131.2 (C-15), 132.6 (C-14), 134.0 (C-14′), 135.2 (C-10′), 135.6 (C-9), 135.6 (C-9′), 135.6 (C-13′), 136.8 (C-13), 136.8 (C-6′), 136.9 (C-8), 137.2 (C-12), 139.4 (C-12′), 142.5 (C-8′), 169.7 (C-4′) (Figure S4).
2.9.3. 5‑Glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al (
3)
HR-ESI-MS (+): *m*/*z* 617.34744 (C
36H
50O
7Na; cald. for 617.34542, ∆ 3.27 ppm). UV-Vis λmax (ε) in MeOH 254 (9200), 315 (6800), 407 (43000), 428 (62000), 452 (56000).
1H NMR (CD3OD) δ: 1.26 (3H, s, H-4), 1.26 (3H, s, H-18), 1.87 (3H, s, H-18′), 1.92 (3H, s, H-19), 1.97 (3H, s, H-20), 1.99 (3H, s, H-20′), 2.03 (3H, s, H-19′), 2.42 (2H, d, *J* =7.5 Hz, H-6), 3.15 (1H, dd, *J* =7.6, 9.0 Hz, H-2″), 3.24 (1H, m, H-5″), 3.26 (1H, m, H-4″), 3.35 (1H, dd, *J* =8.9, 9.0 Hz, H-3″), 3.65 (1H, m, H-6″a), 3.82 (1H, m, H-6″b), 4.51 (1H, d, *J* =7.6 Hz, H-1″), 5.91 (1H, m, H-7), 6.11 (1H, d, *J* =12.0 Hz, H-10), 6.18 (1H, d, *J* =15.2 Hz, H-8), 6.26 (2H, H-14 and H-14′), 6.37 (1H, d, *J* =15.0 Hz, H-12), 6.51 (1H, d, *J* =12.3 Hz, H-10′), 6.56 (1H, d, *J* =15.0 Hz, H-12′), 6.65 (2H, H-15 and H-15′), 6.68 (1H, dd, *J* =12.0, 15.0 Hz, H-11), 6.72 (1H, dd, *J* =12.3, 14.9 Hz, H-11′), 6.74 (1H, dd, *J* =10.8, 14.9 Hz, H-7′), 6.88 (1H, d, *J* =14.9 Hz, H-8′), 7.10 (1H, d, *J* =10.8 Hz, H-6′), 9.39 (1H, s, H-4′) (Figure S5).
13C NMR (CD
3OD) δ: 8.0 (C-18′), 11.2 (C-20), 11.3 (C-19′)
c, 11.5 (C-20′)
c, 11.6 (C-19), 25.3 (C-18), 25.6 (C-4), 29.3 (C-18), 45.3 (C-6), 61.4 (C-6″), 70.4 (C-4″), 73.8 (C-2″), 76.2 (C-5″), 76.8 (C-3″), 77.8 (C-5), 97.3 (C-1″), 122.1 (C-7′), 123.0 (C-11′), 123.7 (C-11), 125.6 (C-7), 129.7 (C-15′)
b, 130.3 (C-10), 131.2 (C-15)
b, 132.2 (C-14)
a, 134.8 (C-9′), 134.9 (C-14′)
a, 135.4 (C-9), 135.8 (C-13′), 135.9 (C-13), 136.1 (C-5′), 137.0 (C-12), 137.4 (C-8), 137.9 (C-10′), 141.0 (C-12′), 146.7 (C-8′), 150.4 (C-6′), 195.1 (C-4′) (Figure S6).
a,b,c: Interchangeable.
2.9.4. 5‑Glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-oic Acid (
4)
HR-ESI-MS (+): *m*/*z* 633.33807 (C
36H
50O
8Na; cald. for 633.34034, ∆ 3.58 ppm). UV-Vis λmax (ε) in MeOH 254 (9200), 315 (6800), 407 (43000), 428 (62000), 452 (56000). 1H NMR (DMSO-*d
6*) δ: 1.14 (6H, s, H-4 and H-18), 1.88 (3H, s, H-19), 1.90 (3H, s, H-18′), 1.93 (3H, s, H-20′), 1.94 (3H, s, H-20), 1.97 (3H, s, H-19′), 2.32 (2H, t, *J* =7.5 Hz, H-6), 2.89 (1H, dd, *J* =7.6, 8.4 Hz, H-2″), 3.02 (1H, dd, *J* =8.4, 8.4 Hz, H-4″), 3.07 (1H, dd, *J* =5.2, 8.4 Hz, H-5″), 3.14 (1H, dd, *J* =8.4, 8.4 Hz, H-3″), 3.39 (1H, dd, *J* =5.2, 11.0 Hz, H-6″b), 3.62 (1H, d, *J* =11.0, H-6″a), 4.32 (1H, d, *J* =7.6 Hz, H-1″), 5.88 (1H, td, *J* = 7.5, 15.6 Hz, H-7), 6.13 (1H, d, *J* = 12.0 Hz, H-10), 6.14 (1H, d, J = 15.6 Hz, H-8), 6.33 (1H, d, *J* = 10.5 Hz, H-14), 6.37 (1H, d, *J*=15.0 Hz, H-12), 6.42 (1H, d, *J* = 10.5 Hz, H-14′), 6.43 (1H, d, *J*=10.8 Hz, H-10′), 6.50 (1H, d, *J*=15.4 Hz, H-12′), 6.55 (1H, dd, *J* = 11.0, 15.0 Hz, H-7′), 6.65 (1H, dd, *J*=12.0, 15.0 Hz, H-11), 6.68 (1H, dd, J=12.2, 15.0 Hz, H-11′), 6.71 (1H, d, *J*=15.0 Hz, H-8′), 6.73 (2H, H-15 and H-15′), 7.18 (1H, d, *J*=11.0 Hz, H-6′) (Figure S7).
13C NMR (DMSO-*d
6*) δ: 12.8 (C-20), 12.8 (C-19′), 12.8 (C-20′), 13.0 (C-18′), 13.1 (C-19), 26.4 (C-4), 26.9 (C-18), 45.1 (C-6), 61.4 (C-6″), 70.4 (C-4″), 73.8 (C-2″), 76.7 (C-5″), 76.9 (C-5), 77.2 (C-3″), 97.4 (C-1″), 123.5 (C-7′), 125.2 (C-11′), 125.7 (C-11), 126.7 (C-5′), 127.1 (C-7), 130.3 (C-10), 130.3 (C-15), 130.3 (C-15′), 132.6 (C-14), 134.2 (C-13′), 135.5 (C-14′), 135.5 (C-9′), 135.8 (C-10′), 136.3 (C-9), 137.0 (C-8), 137.0 (C-13), 137.2 (C-12), 138.3 (C-6′), 139.8 (C-12′), 143.8 (C-8′), 169.5 (C-4′) (Figure S8).
*3.1. Isolation of the crtP and aldH Genes from P. maritimus Strain iso-3*
Previously, we isolated the carotenoid biosynthesis gene cluster from *P. maritimus* iso-3, which contains *crtN* (*crtNa*), *crtM*, *crtNb*, *GT*, and *cruF* [
3]. However, these genes were not sufficient to synthesize the carotenoid of *P. maritimus*, such as methyl 5-glucosyl-5,6-dihydro-4,4′-diapolycopenoate. Recently, Lee et al. cloned *crtP* and *aldH* genes from *P. faecalis* as carotenogenic genes [
13]. Since the genome sequences are similar between the *Planococcus* genera, we designed PCR primers for these genes based on the genome sequences of *P. faecalis* and *P. plakorditis* (Table S1). PCR cloning with these primers resulted in the isolation of two genes from *P. maritimus* iso-3.
In *Staphylococcus aureus*, Pelz et al. found the gene mediating the oxidation of the terminal methyl group in a C
30-carotenoid and named it *crtP* [
8]. The *crtP* designation was adopted for the ortholog in *P. faecalis* [
13]. In contrast, the *crtP* gene has been widely designated as the cyanobacterial phytoene desaturase gene [
14]. Because the same gene name for distinct carotenoid genes is confusing, we propose that *crtP* be renamed *cruO* for the carotenoid terminal oxidase gene.
*3.2. Sequence Analysis of the Carotenoid Biosynthesis Gene Candidates*
Subsequently, we performed a sequence analysis of the *cruO* and *aldH* genes isolated from *P. maritimus* iso-3. The predicted amino acid sequence of the *cruO* gene product (CruO) showed 86% identity to those of the *P. faecalis* CrtP, while CruO was 29%, 25%, and 23 % identical to CrtNb, CrtNa, and Orf2 that were isolated from the same *P. maritimus* iso-3, respectively. The phylogenetic tree of *crtN*-homologous genes () showed that the *P. maritimus* iso-3 *cruO* gene encoding carotenoid terminal oxidase, along with the other corresponding genes (*crtP*), fell into the *crtNb* clade. This clade included the *crtNb* gene encoding carotenoid desaturase from *P. maritimus* iso-3 at earlier branch point (). On the other hand, the predicted amino acid sequences of AldH showed 79% identity to those of the *P. faecalis* AldH. These *aldH* genes were found to belong to another gene family different from the *crtNc* (*aldH*) clade.
. Phylogenetic tree of *crtN*-homologous genes.
Amino acid alignment of the encoded proteins was performed using MAFFT (http://www.mafft.ccbrc.jp/) and phylogenetic trees was constructed using the neighbor joining method, both as implemented on the GenomeNet (https://www.genome.jp/tools/ete/). Accession numbers of the sequences are listed in .
. Examples of crtN-homologous genes and their accession numbers.
*3.3. CruO Acts as an Aldehyde Synthase*
To investigate the function of the *cruO* gene from *P. maritimus* iso-3, we constructed plasmids pAC-HIMNFNbP and pAC-HIMNFNbGP () and introduced them into *E. coli* (JM101(DE3)). Recombinant *E. coli* strains carrying plasmids pAC-HIMNFNb and pAC-HIMNFNbG produced 5-hydroxy-5,6-dihydro-apo-4,4′-lycopene (Peak A) and 5-glucosyl-5,6-dihydro-apo-4,4′-lycopene (Peak B), respectively (a,c), as shown previously [
3]. However, new peaks 1 and 3 were observed in *E. coli* carrying pAC-HIMNFNbP and pAC-HIMNFNbGP, respectively (a,c). The produced compounds
1 and
3 were purified and analyzed using ESI-MS (+),
1H, and
13C NMR, as described later. Consequently,
1 and
3 were identified as 5‑hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al and 5‑glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al, respectively. Thus, the *cruO* gene was confirmed to encode the carotenoid terminal oxidase, which converts the terminal methyl group of a C
30-carotenoid into its aldehydes. In the *crtNb* clade (), the Methylomonas 16a *crtNb* gene and the *crtP* genes of *S. aureus* and *P. faecalis* are considered to encode enzymes with similar oxidation activity to CruO [
15].
. Functional analysis of *cruO* (*crtP*) and *aldH* of *P. maritimus* strain iso-3. (a) HPLC chromatograms of the extracts of *E. coli* transformants carrying pAC-HIMNFNbP; (b) pAC-HIMNFNbPA; (c) pAC-HIMNFNbGP; and (d) pAC-HIMNFNbGPA. The UV-visible spectra of compounds 1–4 are presented on the right of the chromatograms. Peak A, 5-hydroxy-5,6-dihydro-apo-4,4′-lycopene; peak B, 5-glucosyl-5,6-dihydro-apo-4,4′-lycopene.
*3.4. AldH Acts as an Aldehyde Dehydrogenase*
The *aldH* gene from *P. maritimus* iso-3 is homologous to *aldH* from *P. faecalis*, which encodes an aldehyde dehydrogenase [
13]. The catalytic activity of the *P. maritimus* AldH protein was examined by constructing the plasmids pAC-HIMNFNbPA and pAC-HIMNFNbGPA (), which were individually introduced into *E. coli* (DE3). These *E. coli* transformants generated new carotenoid peaks
2 and
4, respectively (b,d). Their ESI-MS (+),
1H, and
13C NMR spectral analyses indicated compounds
2 and
4 as 5‑hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-oic acid and 5‑glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-oic acid, respectively. These results show that *aldH* codes for aldehyde dehydrogenase, which converts the terminal aldehyde group of a C
30-carotenoid into its carboxylic acid.
3.5. Structural Determination of the Intermediate Carotenoids (1–4)
The molecular formula of
1 was determined as C
30H
40O
2 using HR-ESI-MS analysis. Analyses of
1H and
13C NMR,
1H–
1H DQF COSY, and HMQC spectra of
1 in CDCl3 showed that the structure of
1 was closely related to that of 4, 4′-diapolycopene, while the signals of two singlet methyls (H-4 (δ
H 1.14) and C-4 (δ
C 29.2)) and (H-18 (δ
H 1.14) and C-18 (δ
C 29.3)), one non-oxygenated sp3 CH
2 (H-6 (δ
H 2.32) and C-6 (δ
C 47.4)), one oxygenated quaternary
13C (C-5 (δ
C 71.0)), and one aldehyde (H-4′ (δ
H 9.45) and C-4′ (δ
C 194.4)) were observed only in
1. The linkages between the observed
1H and
13C signals were analyzed using the HMBC spectrum. In the HMBC spectrum, preservation of the all *trans* olefin structure of 4,4′-diapolycopene from C-7 to C-5 in
1 was proved using the
1H-
13C long-range couplings from the methyl signals of H-19 (δ
H 1.94), H-20 (δ
H 1.98), H-18′ (δ
H 1.90), H-19′ (δ
H 2.02), and H-20′ (δ
H 1.98) (). The
1H-
13C long range couplings from H-4 and H-18 to C-5 and C-6 and vicinal
1H-
1H spin coupling between H-6 and H-7 showed that the 2-methyl, 2-oxygenated propyl structure composed of C-4, C-5, C-6, and C-18 were attached at C-7. Furthermore, the aldehyde function at C-4′ was demonstrated using the
1H-
13C long-range coupling from H-18′ to C-4′. Based on these observations, the structure of
1 was determined to be 5‑hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al (). According to the CAS database, compound
1 was new.
. Structural determination of compounds 1–4.
The molecular formula of
2 was determined to be C
30H
40O
3 by HR-ESI-MS analysis. The
1H and
13C NMR spectra of
2 in CDCl
3 were almost identical to those of
1 except for the disappearance of the aldehyde signal in
1 and the appearance of a carbonyl signal (C-4′ (δ
C 169.7)). Because long-range coupling from H-18′ (δ
H 1.88) to C-4′ was observed in the HMBC spectrum and the molecular formula of
2 was
1 + O, the structure of
2 was determined as 5‑hydroxy‑5,6‑dihydro‑4,4′‑diapolycopen-4′-oic acid (). Compound
2 was a new compound according to the CAS database.
The molecular formula of
3 was determined to be C
36H
50O
7 using HR-ESIMS analysis. The
1H and
13C NMR,
1H–
1H DQF COSY, and HMQC spectra of
3 in CD3OD revealed that the structure of
3 was closely related to
1, while
1H and
13C signals derived from a hexose were observed in
3. Hexose was identified as β-glucose by
1H–
1H vicinal spin couplings of H-1′–H-6′ and the linkage of the glucose at C-5 was shown by the
1H-
13C long-range coupling from H-1″ (δ
H 4.51) to C-5 (δ
C 70.4) observed in the HMBC spectrum. Based on these findings, the structure of
3 was determined to be 5‑glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-al (). According to the CAS database, compound
3 was new.
The molecular formula of
4 was determined to be C
36H
50O
7 using HR-ESI-MS analysis. The
1H,
13C NMR,
1H–
1H DQF COSY, and HMQC spectra of
4 in DMSO-*d
6* revealed that the structure of
4 was almost identical to
3, while the aldehyde signal in
3 disappeared in
4 and a carbonyl signal (C-4′ (δ
C 169.5)) was observed only in
4. Since the long-range coupling from H-18′ (δ
H 1.90) to C-4′ was observed in the HMBC spectrum () and the molecular formula of
4 was compound
3 + O, the structure of
4 was determined as 5‑glucosyl‑5,6‑dihydro‑4,4′‑diapolycopen-4′-oic acid (). According to the CAS database, compound
4 was new.
*3.6. Singlet Oxygen-quenching Activities of Newly Generated Carotenoids*
The singlet oxygen-quenching activities of the newly generated C
30-carotenoids (intermediates) were examined and the results are shown in . As shown in , compounds closer to the end of the biosynthetic pathway exhibited more potent singlet oxygen-quenching activity.
. Singlet oxygen-quenching activity of the individual C30-carotenoids (intermediates).
This result suggests that *P. maritimus* iso-3 living near the surface of the sea produces metyl-5-glucosyl-5,6-dihydro-4,4′-diapolycopenate to protect itself from singlet oxygen damage caused by ultraviolet and triplet oxygen. The dipolar structure of this highly modified C
30-carotenoid may also stabilize the phospholipid double-layer membrane of Planococcus, and consequently may contribute to the tolerance of this bacterium not only to photooxidative damage but also to solvents [
3].
A similar highly modified dipolar C
30-carotenoid, glycosyl-4,4′-diaponeurosporen-4′-ol-4-oate, is found in the same genus *P. faecalis* [
13]. On the other hand, *S. aureus* is known to produce staphyloxanthin as the monopolar C
30-carotenoid, whose biosynthetic pathway has been elucidated at the gene level [
7,
8]. We have elucidated the biosynthetic pathway of highly modified dipolar C
30-carotenoids for the first time, using the *Planococcus* genes.
The following supporting information can be found at: https://www.sciepublish.com/index/journals/article/sbe/25.html/id/17.
Data Availability: Accession numbers of the *cruO* (*crtP*) and *aldH* genes are LC722835 and LC722836, respectively. Accession of the sequences of the plasmids, pAC-HIMNFNbP, pAC-HIMNFNbGP, pAC-HIMNFNbPA and pAC-HIMNFNbGPA are LC722947, LC722948, LC722949, and LC722950, respectively.
Conceptualization: K.S. and N.M.; Investigation: M. H., C.M., and M.T.; Writing – Original Draft Preparation: K.S., M.T., and N.M.
Not applicable.
Not applicable.
This research received no external funding.
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.