1. Introduction
Forest canopies are unique zones of biodiversity [
1]. In tropical rainforests, arthropod abundance and diversity in the upper canopy are 2–4 times greater than those in the understory [
2]. In Sulawesi, the number of beetle species increased by 50% every 10 m in vertical height [
3]. However, some studies have found that several insect groups, including Coleoptera, Hymenoptera, and Lepidoptera, at the ground may be as speciose as at the canopy [
4,
5,
6]. Nevertheless, many arthropod groups show clear patterns of stratification, particularly in complex tropical rainforests [
7,
8], which may be related to different habitat conditions [
1,
9]. In general, arthropod assemblages, including mites (Arachnida) [
10], Collembola [
11], Diptera [
12], Formicidae [
13], Lepidoptera [
14,
15] in the canopy are very distinct from those that inhabit the understory [
16].
Although vertical microclimatic and biotic gradients are much steeper in tropical compared to temperate forests [
17,
18,
19], stratification patterns are similar [
20,
21,
22,
23].
Using flight interception traps, Ulyshen and Hanula [
24] collected approximately 29% and 31% of beetle species exclusively in the canopy or near the ground, respectively, in a temperate deciduous forest in the USA. Similarly, 30% of 88 beetle species were caught only in the canopy and 30% exclusively in the ground layer in an Italian temperate forest [
25].
In tropical rainforests, there are commonly five recognized strata [
9], with the understory often defined as the vegetation layer from the ground cover to the lower canopy [
26]. Still, stratification differs even within and between tropical lowland rainforests, as the number and distinctness of vegetation strata vary significantly in relation to floristic composition [
27,
28]. The understory tree composition of Neotropical and African rainforests consists of many small flowering trees, whereas non-reproductive juveniles of canopy trees dominate in Southeast Asian rainforests [
29]. In Amazonian forests, the dominant plant species in the understory are distinct from those in the upper canopy [
30,
31], as found in the Venezuelan study site [
32]. Within Amazonia, periodically flooded forests (várzeas and igapós) may exhibit contrasting vertical stratification dynamics compared to non-flooded terra firme forests [
33], such as prevailing in the Venezuelan study site.
As canopy and understory differ in terms of resource availability [
34,
35], strata preferences might be mitigated by the use of certain food resources, sometimes changing between seasons [
36,
37,
38]. Guilds, such as scavengers, may occur predominantly in the understory, whereas many herbivores inhabit the upper canopy [
7]. Otherwise, the proportion of shared gallers and miners in both the canopy and understory was only 6% in a Panamanian wet forest [
39]. Even the same food resources can attract different species in diverse strata. Barrios [
40] compared herbivorous species on saplings and trees and found that only one leaf-chewing chrysomelid species occurred on both. Similarly, the composition of flower-visitor communities generally exhibits large differences between the canopy and understory in tropical [
41,
42] and temperate forests [
43].
Studying vertical stratification is important because it is often considered a key factor in promoting extreme diversity in tropical forests [
44] through niche partitioning among different functional guilds [
45]. Here, I analyze the occurrence of 862 canopy beetle species from 45 families collected from 23 Amazonian tree species in the lower understory to characterize their stratum associations. I hypothesize that strata use is related to adult food resources and larval substrates. Specifically, predatory or omnivore beetle taxa should comprise a high proportion of stratum generalists in comparison to phytophagous families. Among the phytophagous families, particularly taxa with larvae developing in the soil should be represented with many species in the lower understory. This study represents the first ecological evaluation of the strata use of a comprehensive canopy beetle assemblage in a tropical rainforest. Furthermore, I compared the diel activity of adult beetles between both strata. A previous analysis of the canopy beetle community revealed that consumers of flowers and extrafloral nectaries visit their host trees either during the day or night [
46]. A stratum switch between the day and night might be indicated if adult beetles show different activity periods in the canopy and lower understory.
2. Materials and Methods
2.1. Study Site
The Venezuelan study site is part of the Man and Biosphere Reserve Alto Orinoco-Casiquiare, which is located in the state of Amazonas (3°10′ N, 65°40′ W; 105 m asl). A canopy crane was installed at the small black-water river Surumoni, a tributary of the Orinoco. This mobile tower crane allowed access to the tree crowns in an area of about 1.4 ha.
The annual precipitation in the study area is approximately 3100 mm [
47]. There is no distinct dry season, but the rainfall reaches the highest values from May to July, showing a lower peak in September and October. Monthly precipitation is at least 100 mm, and air humidity is between 85–90% [
48]. The average annual temperature in the study area is approximately 26 °C with maximum temperatures of up to 30.5 °C during the day and only 20–21 °C during the night.
2.2. Vegetation
The lowlands of the study site belong to the Imerí province [
49], which covers a large area from Brazil to southern Venezuela, Colombia, and Peru. The remote area is covered by lowland moist rainforest. The forest in the study area is frequently interrupted by light gaps owing to irregular crown closures. The upper canopy usually ranges from 25 to 27 m in height.
Altogether, 322 species of spermatophyta were identified in the 1.4 ha crane plot, belonging to 208 genera from 78 families [
32]. Frequent tree species within the crane plot included
Couma utilis (Mart.) Müll. Arg. (Apocynaceae),
Dialium guianense (Aubl.) Sandwith (Fabaceae),
Goupia glabra Aubl. (Goupiaceae),
Ocotea aff.
amazonica (Meisn.) Mez (Lauraceae),
Oenocarpus bacaba Mart. (Arecaceae),
Podocalyx loranthoides Klotzsch (Picrodendraceae), and
Ruizterania trichanthera (Spruce ex Warm.) Marc.-Berti (Vochysiaceae).
The underlayer growth (5–10 m) consisted of young
G. glabra and
O. bacaba [
32]. The herb layer was well-developed. It was dominated by ferns of the families Hymenophyllaceae and Metaxyaceae, as well as small palms of the genera
Geonoma Willd. and
Bactris Jacq. ex Scop. Other abundant plants included Rubiaceae (
Psychotria L. and
Faramea Aubl.), Melastomataceae, Maranthaceae (
Ischnosiphon Körn.), and Heliconiaceae. The forest floor was sparsely covered with leaf litter.
2.3. Field Studies
The general beetle survey was conducted as part of an interdisciplinary research project organized by the Austrian Academy of Science. The field study was carried out between 1997 and 1999 and covers a cumulative year. The samples in the canopy and understory of the crane plot were obtained during the same three periods but differ slightly because of different canopy access: Canopy: September–November 1997, May–August and December 1998, and January–April 1999; Understory: August–November 1997, April–August and December 1998, and January–March 1999. Permits were issued under the number 15-1277 by Servicio Autonoma de Fauna, Ministerio del Ambiente y de los Recursos Naturales Renovables, Caracas, Venezuela.
To evaluate stratum associations of adult beetles, imagines were observed and collected in both the lower understory and canopy every second day and night. Both strata samples combine hand and trap sampling so that the methods complement each other by accessing the species inventory [
50,
51,
52,
53].
The canopy beetle survey included 23 tree species representing 13 plant families, which were monitored throughout the cumulative year and sampled with flight interception traps. The description of the selected trees and sampling protocols were provided separately [
50]. Although, the beetle assemblage comprised only a small proportion of the potential beetle diversity, it is representative of the 23 tree species [
50]. Due to the different plant composition between the canopy and understory, only the fruits of two out of 23 tree species were collected in the two strata. Moreover, the canopy focused on adult beetle host associations comprising leaves, flowers, and fruits, whereas understory samples included the ground, trunks, herbs, and fruit falls.
In the understory, pitfall traps (three traps per site; diameter: 6 cm) were used to collect ground-dwelling beetles. The traps were filled to one-third of their height with water mixed with a surface-tension-diminishing detergent. The beetles were extracted two or three times per week. Pitfall traps were installed at available fruit falls, as such resource concentrations are known to attract beetles [
54,
55]:
G. glabra (all periods),
Miconia poeppigii Triana (Melastomataceae) (December 1998 to March 1999),
O. bacaba (May 1998),
Ficus L. sp. (Moraceae) (September 1997),
Protium cf.
spruceanum (Benth.) Engl. (Burseraceae) (August–September 1997; May–August 1998),
Alibertia cf.
latifolia (Benth.) K. Schum. (Rubiaceae) (March 1999; July 1998), and
Esenbeckia Kunth sp. (Rutaceae) (August–September 1997). Hand collection was conducted around the fruit falls and along a 40 m long transect on a small path in the crane plot. Herbs, young trees, and trunks up to a height of approximately 1.5 m and the ground were checked visually for adult beetles and manually collected.
Manual collection is considered to be equivalent in both strata, but was not structured to provide quantitative data. In contrast, the traps obtained (semi-)quantitative data. However, the trap samples may have caused a strong bias because the canopy window traps collected flying beetles [
51], and understory trapping focused on ground-dwelling beetles [
56]. In particular, the predominantly predatory Carabidae are well represented in the pitfall traps [
57,
58]. As the traps focused largely on different guilds, the number of shared beetle species may be lower than expected using identical methods.
2.4. Beetle Characterization and Identification
To identify factors underlying beetle host tree associations, adult canopy beetles were ecologically characterized. To test my hypothesis that the availability of food resources may influence the strata’s use of adult beetles, I assigned host tree identity and diet to the beetle species. Host plants include all canopy tree species where the beetles were collected. The diet (leaves, flowers, extrafloral nectar, fruits) was either directly observed or beetles exclusively associated with a phenological season of the trees were assigned as flower or extrafloral nectary visitors. As diel activity may differ between the forest strata, I monitored the activity of adult beetles in both strata. Diel activity was distinguished as nocturnal or diurnal if all individuals of one species showed uniform activity either during the day or night. The activity was observed for about two minutes and comprised all behaviors (walking, flying, feeding, mating, or cleaning) except for quiescent resting.
The collected beetles were preserved in 70% ethanol and later, in part, pinned. They were assigned to morphospecies (hereafter species), and species of the families Cantharidae, Chrysomelidae, Curculionidae, Elateridae, Lampyridae, Scarabaeidae, and Tenebrionidae were identified by experts [
59]. Carabidae were identified by the author using Reichardt [
60]. The higher classification follows Bouchard et al. [
61]. Voucher specimens were deposited at the Museo del Instituto de Zoología Agrícola ‘Francisco Fernandez Yepez’, Maracay, Venezuela.
2.5. Data Processing and Analyses
Every adult beetle collected from the 23 target tree species was included in the dataset. The complete list of beetle families, including their species numbers and abundances and a dataset comprising all canopy species with their host trees were published previously [
59,
62]. In the first step, all individuals of canopy beetle species collected from the ground/understory were added to this dataset (Table
1). In my analyses, I consider all beetle families represented with at least 20 species at the canopy and all beetle families containing species sampled in both the canopy and lower understory (a). Subsequently, all the unique singletons were omitted from the dataset for a tighter evaluation (b). Third, all congenerics collected from the ground/understory were added to the identified canopy genera (Table
2). Whereas all species of Cantharidae, Carabidae, Elateridae, Lampyridae, and Scarabaeidae collected in the understory were assigned to a genus, the genus identification of Chrysomelidae and Curculionidae was carried out only in part. Therefore, the total number of species per genus in Chrysomelidae and Curculionidae may be larger than that given in Table
2.
To evaluate if diel activity of beetle species differs between the canopy and understory, I compared the diel activity of observed beetle species between both strata. Furthermore, I compared the attraction of beetles to host plant resources between both strata to evaluate if the same resources attract the same beetles.
Although the samples in both strata were obtained in the same study plot during the same periods, I consider the canopy and the understory samples as independent due to the following reasons: the plant species and resources, respectively, differed largely between both strata and the trap samples focused on different beetle activity.
However, based on similar sampling protocols, a good coverage of the inventory [
50], but overall low abundances, I combined trap and hand samples for each stratum to compare the distribution of species abundances, as well as the number of species and genera of selected taxa shared between the canopy and understory.
. Canopy beetle families collected with at least 20 species and/or comprising species shared between canopy and lower understory collected from 23 canopy tree species and in the understory in a lowland tropical rainforest in Venezuela for a cumulative year between 1997 and 1999: (<b>a</b>) total number of species; (<b>b</b>) number of species excluding singletons.
To analyze the statistical significance of the differences between canopy and understory samples, the non-parametric Mann-Whitney U test was used. This test compares the medians of two independent samples and assumes an equal-shaped distribution rather than a normal distribution for both groups. Analyses were performed using PAST (Version 4.17) [
63].
3. Results
3.1. Shared Species: Overview
In total, 862 adult beetle (morpho-)species from 45 families were collected from 23 canopy tree species during the cumulative year. Of these, 70 species (8.1%) from 11 families were found in the herb layer and/or on the ground (). Excluding all singletons from the cumulative sample (s = 480), 14.6% of all species were shared between the canopy and lower understory.
The majority of the 70 species (62.9%, s = 44) were sampled with more individuals in the canopy, while 15 species (21.4%) were collected with more individuals in the understory (
U = 1381,
p < 0.001). Eleven species (15.7%) were represented by equal proportions of canopy and understory individuals. Sixteen (22.6%) of the 70 shared species were represented as single individuals in the canopy sample. In contrast, 39 shared species (55.7%) were collected as single specimens in the understory.
.
Adult beetle species collected from 23 canopy tree species and in the understory in a lowland tropical rainforest in Venezuela for a cumulative year between 1997 and 1999.
Family (Subfamily)
Species |
Total Individuals |
Canopy Individuals |
Understory Individuals |
Canopy Activity |
Understory Activity |
| Cantharidae (Silinae) |
|
|
|
|
|
| Ditemnus LeConte sp. 1 |
6 |
4 |
2 |
|
day |
| Carabidae (Hapalinae) |
|
|
|
|
|
| Agra Fabricius sp. 4 |
2 |
1 |
1 |
night |
day |
| Apenes LeConte sp. 1 |
5 |
4 |
1 |
|
|
| Apenes LeConte sp. 2 |
14 |
4 |
10 |
|
day/night |
| Lebia Latreille cf. soror |
3 |
2 |
1 |
|
night |
| Loxandrus cf. tetrastigma Bates |
9 |
1 |
8 |
|
night |
| Loxandrus LeConte sp. 2 |
29 |
8 |
21 |
night |
day/night |
| Selenophorus Dejean cf. discopunctatus |
4 |
1 |
3 |
day |
day |
| Notiobia flavicinctus Erichson |
5 |
1 |
4 |
|
night |
| Notiobia nebrioides Perty |
207 |
5 |
202 |
|
night |
| Pentagonica Schmidt-Goebel sp. 1 |
3 |
2 |
1 |
night |
|
| Carabidae (Scaritinae) |
|
|
|
|
|
| Ardistomis cf. fasciolatus Putzeys |
7 |
6 |
1 |
night |
|
| Clivina Latreille sp. 1 |
7 |
3 |
4 |
night |
|
| Clivinina sp. 3 |
13 |
12 |
1 |
night |
|
| Solenogenys funkei Adis |
3 |
2 |
1 |
night |
day |
| Stratiotes Putzeys sp. 1 |
17 |
2 |
15 |
|
|
| Chrysomelidae (Alticini) |
|
|
|
|
|
| Hypolampsis Clark sp. 4 |
2 |
1 |
1 |
|
|
| Omophoita aequinoctialis (Linné) |
6 |
1 |
5 |
night |
day |
| Chrysomelidae (Galerucinae) |
|
|
|
|
|
| Diabrotica cf. alboplagiata Jacoby |
9 |
8 |
1 |
night |
day |
| Diabrotica cf. septemliturata Erichson |
41 |
40 |
1 |
night |
day |
| Diabrotica Chevrolat sp. 4 |
4 |
3 |
1 |
night |
day |
| Diabrotica Chevrolat sp. 6 |
5 |
4 |
1 |
night |
day |
| Diabrotica Chevrolat sp. 7 |
121 |
119 |
2 |
night |
day/night |
| Diabrotica Chevrolat sp. 9 |
15 |
14 |
1 |
night |
day |
| Zepherina cf. variegatus (Weise) |
38 |
36 |
2 |
crepuscular |
day |
| Chrysomelidae (Eumolpinae) |
|
|
|
|
|
| Antitypona cf. lateralis Baly |
5 |
4 |
1 |
day/night |
|
| Brachypnoea Gistel sp. |
17 |
5 |
12 |
day/night |
day |
| Colaspis Fabricius sp. 1 |
24 |
23 |
1 |
day/night |
night |
| Colaspis callichloris Lefévre |
2 |
1 |
1 |
night |
day |
| Entomochirus cf. hirtus |
2 |
1 |
1 |
|
|
| Sphaeropis Lefévre sp. |
5 |
4 |
1 |
|
|
| Spintherophyta Dejean sp. 1 |
14 |
13 |
1 |
day/night |
|
| Curculionidae (Baridinae) |
|
|
|
|
|
| Baridinae sp. 4 |
5 |
3 |
2 |
|
day |
| Baridinae sp. 49 |
118 |
117 |
1 |
day/night |
day |
| Baridinae sp. 29 |
17 |
13 |
4 |
day |
day |
| Curculionidae (Cryptorhynchinae) |
|
|
|
|
|
| Cryptorhynchinae sp. 1 |
4 |
3 |
1 |
|
|
| Cryptorhynchinae sp. 28 |
2 |
1 |
1 |
|
day |
| Curculionidae (Entiminae) |
|
|
|
|
|
| Compsus cf. albus Hustache |
21 |
18 |
3 |
night |
day/night |
| Naupactus Dejean sp. |
57 |
56 |
1 |
day/night |
day |
| Curculionidae (Molytinae) |
|
|
|
|
|
| Conotrachelini sp. 1 |
3 |
2 |
1 |
|
|
| Conotrachelini sp. 7 |
2 |
1 |
1 |
night |
night |
| Cleogonus marginesulcatus Chevrolat |
23 |
21 |
2 |
night |
day |
| Heilipodus Kuschel sp. 1 |
2 |
1 |
1 |
|
day |
| Heilipodus Kuschel sp. 6 |
45 |
44 |
1 |
night |
day |
| Heilipodus Kuschel sp. 8 |
31 |
30 |
1 |
night |
day |
| Heilipodus albobrunneus Rheinheimer |
33 |
29 |
4 |
day/night |
day |
| Heilipodus suspensus (Pascoe) |
5 |
2 |
3 |
night |
day |
| Heilus Kuschel sp. 1 |
99 |
95 |
4 |
night |
day |
| Hilipinus Champion sp. |
5 |
4 |
1 |
night |
day |
| Homalinotus kuscheli Vaurie |
7 |
6 |
1 |
day/night |
|
| Ozopherus muricatus Pascoe |
4 |
3 |
1 |
day/night |
day |
| Curculionidae (Rhynchophorinae) |
|
|
|
|
|
| Metamasius hemipterus (Linné) |
8 |
4 |
4 |
|
day |
| Elateridae |
|
|
|
|
|
| Chalcolepidius limbatus (Fabricius) |
8 |
6 |
2 |
night |
day |
| Crepidius ophthalmicus Candèze |
3 |
2 |
1 |
night |
day |
| Dipropus Germar sp. 3 |
3 |
1 |
2 |
night |
night |
| Semiotus ligneus (Linnaeus) |
13 |
10 |
1 |
night |
day |
| Lampyridae |
|
|
|
|
|
| Cratomorphus Motschulsky sp. 1 |
20 |
16 |
4 |
night |
night |
| Nitidulidae |
|
|
|
|
|
| Nitidulidae sp. 8 |
8 |
4 |
4 |
|
|
| Nitidulidae sp. 9 |
7 |
4 |
3 |
|
|
| Nitidulidae sp. 10 |
42 |
1 |
41 |
|
|
| Ptilodactylidae |
|
|
|
|
|
| Ptilodactylidae sp. 2 |
15 |
14 |
1 |
|
|
| Scarabaeidae (Cetoniinae) |
|
|
|
|
|
| Hoplopyga liturata (Olivier) |
7 |
1 |
6 |
day |
day/night |
| Scarabaeidae (Dynastinae) |
|
|
|
|
|
| Stenocrates cf. carbo Prell |
6 |
4 |
2 |
night |
day |
| Scarabaeidae (Melolonthinae) |
|
|
|
|
|
| Barybas Blanchard sp. 3 |
12 |
11 |
1 |
night |
day |
| Scarabaeidae (Rutelinae) |
|
|
|
|
|
| Cnemida retusa (Fabricius) |
46 |
45 |
1 |
day |
night |
| Leucothyreus MacLeay sp. |
4 |
2 |
2 |
night |
day |
| Macraspis festiva Burmeister |
32 |
31 |
1 |
day |
day |
| Macraspis MacLeay sp. 1 |
2 |
1 |
1 |
|
day |
| Tenebrionidae |
|
|
|
|
|
| Uleda diaperoides Laporte de Castelnau |
5 |
1 |
4 |
night |
|
| Zopheridae |
|
|
|
|
|
| Monommatini sp. 2 |
22 |
21 |
1 |
night |
night |
3.2. Shared Species: Families
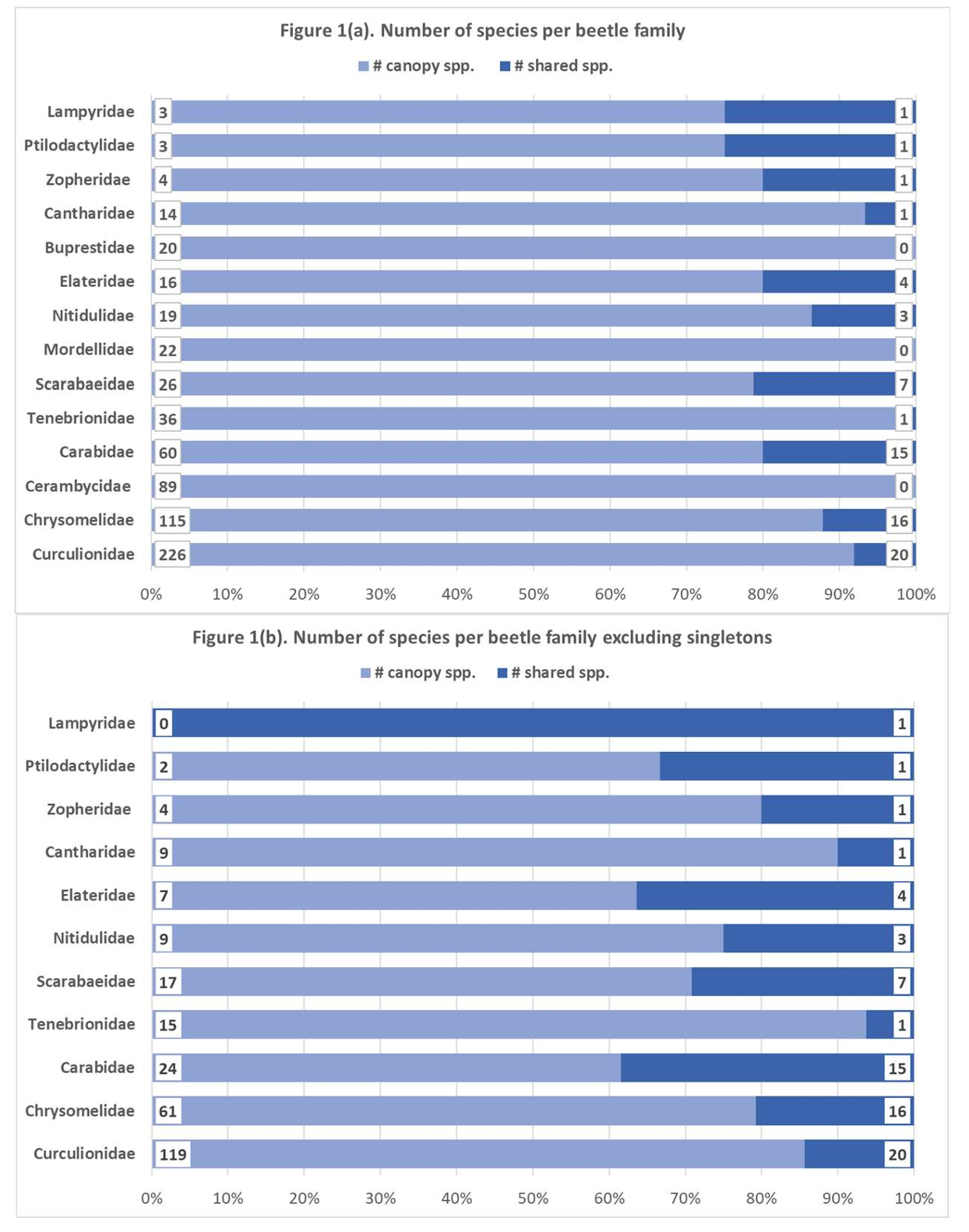
Most canopy species represented in the understory sample were Curculionidae (s = 20), Chrysomelidae (s = 16), and Carabidae (s = 15), followed by Scarabaeidae (s = 7), Elateridae (s = 4), and Nitidulidae (s = 3) (
). The families Cantharidae, Lampyridae, Ptilodactylidae, Tenebrionidae, and Zopheridae comprised only one species represented in both strata.
Among the ten most species-rich beetle families in the canopy (s ≥ 20), Buprestidae, Cerambycidae, and Mordellidae were not recorded in the understory (
a). According to the number of species collected in the canopy, Scarabaeidae (21.2%), Carabidae (20%), and Elateridae (20%) had the highest proportions of shared species. Excluding all singletons (
b), 38.5% of Carabidae, 36% of Elateridae, and 29.2% of Scarabaeidae were shared between the two strata.
Among Curculionidae, most shared species (s = 15 (75%);
U = 75,
p = 0.0005) were collected with more individuals in the canopy (
). The same applies to shared Chrysomelidae species (s = 11 (68.75%);
U = 50.5,
p = 0.0019). In contrast, most shared carabid species (s = 8 (53.3%);
U = 110,
p = 0.93) were represented by more individuals in the understory, but this was not statistically significant.
3.3. Genera Distribution
Forty-four of the 70 shared species were assigned to 29 genera sampled, with at least two species in the crane plot (
). These 29 genera in seven families comprised 145 species, with 130 species (89.7%) collected in the canopy and 59 species (40.7%) collected in the understory (
U = 217,
p = 0.0005). Six out of ten carabid genera (60%;
U = 44.5,
p = 0.69) were collected with more species in the understory, but the difference was not significant. Except for the eumolpine genus
Colaspis Fabricius, the genera of all other beetle families had more representatives in the canopy than in the understory.
.
Number of species in beetle genera (≥2 spp.) collected from 23 canopy tree species and in the understory in a lowland tropical rainforest in Venezuela for a cumulative year between 1997 and 1999.
| Family |
Genus |
Total
spp. |
Canopy
spp. |
Understory
spp. |
# Shared
spp. |
| Cantharidae |
Ditemnus LeConte |
3 |
3 |
1 |
1 |
| Carabidae |
Agra Fabricius |
31 |
31 |
1 |
1 |
| Carabidae |
Apenes LeConte |
4 |
2 |
4 |
2 |
| Carabidae |
Lebia Latreille |
7 |
7 |
1 |
1 |
| Carabidae |
Loxandrus LeConte |
3 |
2 |
3 |
2 |
| Carabidae |
Notiobia Perty |
5 |
2 |
5 |
2 |
| Carabidae |
Pentagonica Schmidt-Goebel |
2 |
2 |
1 |
1 |
| Carabidae |
Selenophorus Dejean |
2 |
1 |
2 |
1 |
| Carabidae |
Ardistomis Putzeys |
2 |
2 |
1 |
1 |
| Carabidae |
Clivinina Rafinesque |
7 |
3 |
6 |
2 |
| Carabidae |
Stratiotes Putzeys |
2 |
1 |
2 |
1 |
| Chrysomelidae |
Hypolampsis Clark |
4 |
4 |
1 |
1 |
| Chrysomelidae |
Omophoita Chevrolat |
3 |
3 |
1 |
1 |
| Chrysomelidae |
Diabrotica Chevrolat |
16 |
16 |
6 |
6 |
| Chrysomelidae |
Zepherina Bechyné |
3 |
3 |
1 |
1 |
| Chrysomelidae |
Antitypona Weise |
3 |
3 |
1 |
1 |
| Chrysomelidae |
Colaspis Fabricius |
2 |
2 |
2 |
2 |
| Chrysomelidae |
Entomochirus Lefévre |
2 |
2 |
1 |
1 |
| Chrysomelidae |
Spintherophyta Dejean |
3 |
3 |
1 |
1 |
| Curculionidae |
Heilipodus Kuschel |
9 |
8 |
6 |
5 |
| Curculionidae |
Heilus Kuschel |
5 |
5 |
1 |
1 |
| Curculionidae |
Homalinotus |
3 |
3 |
1 |
1 |
| Elateridae |
Crepidius Candèze |
2 |
2 |
1 |
1 |
| Elateridae |
Dipropus Germar |
3 |
3 |
1 |
1 |
| Lampyridae |
Cratomorphus Motschulsky |
2 |
2 |
1 |
1 |
| Scarabaeidae |
Stenocrates Burmeister |
4 |
3 |
2 |
1 |
| Scarabaeidae |
Barybas Blanchard |
3 |
3 |
1 |
1 |
| Scarabaeidae |
Cnemida Kirby |
2 |
2 |
1 |
1 |
| Scarabaeidae |
Macraspis MacLeay |
8 |
7 |
3 |
2 |
| TOTAL spp. |
|
145 |
130 |
59 |
44 |
3.4. Diel Activity
Forty beetle species were observed in both forest strata (
). Twenty-two of these species (55%) were found during different periods of the day in the canopy and understory, whereas seven species were observed during the same period in both strata (
U = 335,
p < 0.0001). Of the 22 species with different diel periods in the canopy and understory, 21 were active in the canopy during the night.
3.5. Resource Use
Ten of 70 beetle species (14.3%) were sampled from the same plants in both the canopy and understory. Seven species, including five species of Carabidae (two species of
Apenes LeConte, two species of
Loxandrus LeConte, and
Notiobia nebrioides Perty) and two species of Nitidulidae were sampled from tree crowns and fruit falls of
G. glabra.
Metamasius hemipterus (Linné) and Baridinae sp. 29 were associated with
O. bacaba in both the strata.
Heilipodus Kuschel sp. 1 was collected from the crown and understory of
C. utilis.
4. Discussion
The majority of beetle species collected from 23 tree species were recorded only at the canopy stratum. While some species-rich families (at least 20 canopy species) were not represented in the lower understory, other species-rich families revealed a proportion of about 20% of species shared between the lower understory and canopy.
In accordance with my hypothesis that the occurrence of canopy beetles in the understory may be related to their ecological characteristics, I found that particularly beetle families with larvae developing in the soil (Elateridae and Scarabaeidae) and predatory Carabidae comprised a high proportion of stratum generalists.
Furthermore, the diel activity of many observed species was different between both strata. Therefore, I attribute this observation to a possible stratum switch between canopy and understory.
4.1. General Stratification
Most beetle species in the sample (85.4%, excluding singletons) were exclusively recorded in the canopy. In contrast, other studies revealed only a few canopy specialists in tropical forests. For instance, Stork and Grimbacher [
6] found 72% of the species (excluding singletons and doubletons) in both strata, with 24 and 27% of the abundant species specializing in the canopy and ground strata, respectively. An Indonesian tropical forest study revealed that only 8–13% of beetles were canopy specialists [
64]. In Sulawesi, 39% of the more abundant nocturnal flying beetle species were specific to the canopy and 5% to the ground layer [
65]. The relatively low percentage of canopy specialists in these studies may be largely caused by the exclusion of rare species, the collection of beetles in mid-layer strata, and the use of light traps that attract beetles from a distance.
The high specificity of the Venezuelan beetle assemblage to the canopy stratum may depend on the distance between both samples. I compared the strata use of adult beetles between the two ends of the vertical gradient at a distance of approximately 20 m. The analyzed beetle assemblage was obtained from the two uppermost strata (18–30 m) and the lower understory up to a height of about 1.5 m. McCaig et al. [
66] suggested that the best fit for vertical stratification is either the distance from the ground or the distance from the canopy. The proportion of shared species versus stratum specialists might not only strongly depend on the exact vertical position of the traps in the forest but also on the methods used to collect beetles, the inclusion criteria for rare species, and the structure of the forests.
4.2. Strata Use and Food Resources
The plant composition and, therefore, the resource availability in the understory were different from those in the canopy in the Venezuelan crane plot. This may have contributed to the high stratum specificity obtained in the canopy beetle assemblage. There were no shared species in Buprestidae, Cerambycidae, or Mordellidae, although these families were sampled with at least 20 species in the canopy. All Mordellidae and most species of Cerambycidae have been exclusively collected from flowering trees [
67]. Buprestidae were either flower visitors or leaf feeders in the canopy (personal observations).
In contrast, Chrysomelidae, Curculionidae, Elateridae, and Scarabaeidae comprised 14.4–36.4% of shared species between both sampled strata in the crane plot. The percentage of 14.4% in Curculionidae was lower in the Venezuelan sample than in a study with light traps in Panama, where no more than 20% of curculionids were restricted to either the canopy or the ground stratum [
68]. Hammond et al. [
64] identified 77 of 140 weevil species as canopy specialists in a Sulawesi lowland rainforest. Yet, 20.8% of the shared leaf-beetle species were in the range of the proportion of chrysomelid species in Panama, where 16% and 28% were shared between the canopy and understory of wet and dry sites, respectively [
69].
Still, the occurrence of some shared beetle species coincided with resource availability on fruit falls of
G. glabra and
O. bacaba on the forest floor. Two species of Nitidulidae and five species of Carabidae, including the seed-feeding species of
Notiobia [
54], were collected on
G. glabra in both strata. Many carabid beetle adults live in soil, leaf litter, or are active on the ground surface [
70]. However, as they are commonly omnivorous or carnivorous [
70], they should be able to forage in different strata of the forest. This may explain why many harpaline and scaritine genera revealed more species inhabiting the forest floor than the canopy, indicating their opportunistic strata use.
4.3. Strata Use and Larval Substrates
Species-rich families represented exclusively in the canopy, including Buprestidae, Cerambycidae, and Mordellidae can complete their entire life cycles in the canopy. Most larvae of these three families feed internally in the stems and leaves of plants or dead wood [
71,
72,
73]. These larval resources are widely available in the canopy.
The high proportions of shared species in Chrysomelidae, Elateridae, and Scarabaeidae may be attributed to oviposition on the forest floor and larvae hatching from the soil. Many larvae of Chrysomelidae [
74], Scarabaeidae [
75], and some Elateridae [
76] develop in the soil. However, there are differences between the subfamilies. The Venezuelan sample comprises six out of seven shared species in the scarab subfamilies Dynastinae, Melolonthinae, and Rutelinae, with many larvae typically feeding on roots [
75]. Within Chrysomelidae, most shared species belong to Eumolpinae and Galerucinae, with most larvae living in the soil and feeding on roots in tropical rainforests [
74,
77]. In contrast, there were no shared species in the subfamily Cryptocephalinae, even though they had the same high number of species in the canopy, such as Eumolpinae [
78]. Most Cryptocephalinae are phytophagous, living and developing on their host plant [
79,
80].
Most shared species were sampled from Curculionidae in the Venezuelan crane plot. Shared species belong predominantly to Molytinae, with most larvae feeding in dead wood or other dead and decaying plant material [
81]. Two of three Entiminae species with larvae feeding externally on roots in the soil [
81] were shared between the canopy and understory. In contrast, the understory sample did not represent Conoderinae, another species-rich subfamily in the canopy stratum (personal observations). Conoderinae often emerges from branches of the canopy stratum [
82].
4.4. Strata Use and Stratum Switch
Twenty-two shared species were found in the canopy and understory during different periods of the day. The use of different strata according to the phases of activity, with many adult beetle species occurring in the forest canopy during the day and resting at night in the understory, was described by Erwin [
83]. He found that Alleculinae feeds on lichens and mosses on tree trunks at night and spends the day hiding in suspended dry leaves elsewhere in the forest. Indeed, 21 of the 22 species with different diel activity in both strata were active in the canopy during the night. This suggests using the understory during the day is a strategy to overcome the hygrothermal stress in the canopy during daytime [
46].
In total, 74.5% of all canopy beetle species visiting either flowers and/or extrafloral nectaries were restricted in their occurrence on host trees to the time of the day [
46,
59]. This permanent diel migration during the course of the day may increase the proportions of stratum generalists, particularly if beetles are collected during the day and night. This might be supported by Basset et al. [
9], who found that the vertical turnover of arthropods, particularly phytophages and predators, exceeds the horizontal and seasonal turnover in a rainforest in Panama. However, the high proportion of beetle species collected only in the canopy and not in the lower understory indicates that most beetles rest between the canopy and lower understory.
5. Conclusions
Amazonian canopy beetles showed high stratum specificity. This may indicate their specialization in arboreal life. However, strata use was related to the ecological characteristics of the taxa. Species-rich canopy beetle families represented with most species in both the canopy and lower understory comprise many larvae developing in the soil: Chrysomelidae, Curculionidae, Elateridae, and Scarabaeidae. The highest proportion of shared species revealed commonly omnivorous or predatory Carabidae that can forage in each stratum independent from the floristic composition. In contrast, the predominantly flower-visiting beetle families Cerambycidae and Mordellidae, with larvae often developing internally in living plant parts or in dead wood available in the canopy, were collected exclusively from the canopy. Thus, stratum use in the Venezuelan crane plot can often be attributed to resource availability and larval development. To evaluate the adaptation of single taxa to arboreal life and get a better foundation for estimating diversity, future studies on stratification should discriminate between ecologically different groups within taxa.
Acknowledgments
The Austrian Academy of Sciences and colleagues are acknowledged for their support and permission to join the Surumoni Project in Venezuela. I cordially thank Jens Wesenberg for assistance in all botanical matters and field work. I am delighted for the identification of beetles by Joachim Adis (Solenogenys funkei Adis); Erik Arndt (Notiobia Perty); Shawn M. Clark, Wills Flowers, David Furth, and Lev N. Medvedev (Chrysomelidae); Paul J. Johnson (Elateridae); Martin Lillig (Tenebrionidae); Alistair S. Ramsdale (Cantharidae); Brett C. Ratcliffe (Scarabaeidae); Sergio Antonio Vanin (Curculionidae); and Vadim R. Viviani (Lampyridae). I appreciate the four reviewers for their constructive feedback, which helped to improve this paper significantly. Leopoldina, Germany, supported this study through scientific, financial, and logistic means.
Author Contributions
The author is responsible for conceptualization, methodology, analysis, investigation, writing of the manuscript and editing.
Ethics Statement
This article does not present research with ethical considerations. Permit: license and authorization for the beetle survey were issued under the number 15-1277 by Servicio Autonoma de Fauna, Ministerio del Ambiente de los Recusos Naturales Renovables, Caracas, Venezuela.
Informed Consent Statement
Not applicable.
Data Availability Statement
The canopy beetle dataset is available at https://doi.org/10.5061/dryad.kprr4xhc7 [
62]. All other data produced in this study are provided in this manuscript.
Funding
The fieldwork was supported, in part, by grants from the ESF Tropical Canopy Programme and the Stiftung der Deutschen Wirtschaft, Germany. Funding for this study is provided by The Royal Society, London, UK.
Declaration of Competing Interest
The author declares that she has no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1.
Nakamura A, Kitching RL, Cao M, Creedy TJ, Fayle TM, Freiberg M, et al. Forests and their canopies: Achievements and horizons in canopy science.
Trends Ecol. Evol. 2017,
32, 438–451. doi:10.1016/j.tree.2017.02.020.
[Google Scholar]
2.
Basset Y. Invertebrates in the canopy of tropical rain forests: How much do we really know?
Plant Ecol. 2001,
153, 87–107.
[Google Scholar]
3.
Davis AJ, Sutton SL. Estimating species richness through three dimensions.
Sepilok Bull. 2012,
15, 69–83.
[Google Scholar]
4.
Ashton LA, Nakamura A, Basset Y, Burwell CJ, Cao M, Eastwood R, et al. Vertical stratification of moths across elevation and latitude.
J. Biogeogr. 2016,
43, 59–69.
[Google Scholar]
5.
Somavilla A, Moraes Junior RNM, Rafael JA. Is the social wasp fauna in the tree canopy different from the understory? Study of a particular area in the Brazilian Amazon Rainforest.
Sociobiology 2019,
66, 179–185. doi:10.13102/sociobiology.v66i1.3568.
[Google Scholar]
6.
Stork NE, Grimbacher PS. Beetle assemblages from an Australian tropical rainforest show that the canopy and the ground strata contribute equally to biodiversity.
Proc. R. Soc. B 2006,
273, 1969–1975.
[Google Scholar]
7.
Basset Y, Hammond PM, Barrios H, Holloway JD, Miller SE. Vertical stratification of arthropod assemblages. In Arthropods of Tropical Forests. Spatio-Temporal Dynamics and Resource Use in the Canopy; Basset Y, Novotny V, Miller SE, Kitching RL, Eds.; Cambridge University Press: Cambridge, UK, 2003; pp. 17–27.
8.
Stork NE, McBroom J, Gely C, Hamilton A. New approaches narrow global species estimates for beetles, insects, and terrestrial arthropods.
Proc. Natl. Acad. Sci. USA 2015,
112, 7519–7523.
[Google Scholar]
9.
Basset Y, Cizek L, Cuénoud P, Didham RK, Novotny V, Ødegaard F, et al. Arthropod Distribution in a Tropical Rainforest: Tackling a Four Dimensional Puzzle.
PLoS ONE 2015,
10, e0144110. doi:10.1371/journal.pone.0144110.
[Google Scholar]
10.
WaIter DE. Dancing on the head of a pin: mites in the rainforest canopy.
Rec. Aust. Mus. 1995,
52, 49–53.
[Google Scholar]
11.
Rogers DJ, Kitching RL. Vertical stratification of rainforest collembolan (Collembola: Insecta) assemblages: Description of ecological patterns and hypotheses concerning their generation.
Ecography 1998,
21, 392–400.
[Google Scholar]
12.
Amorim DS, Brown BV, Boscolo D, Ale-Rocha R, Alvarez-Garcia DM, Balbi MIPA, et al. Vertical stratification of insect abundance and species richness in an Amazonian tropical forest
. Sci. Rep. 2022,
12, 1734. doi:10.1038/s41598-022-05677-y.
[Google Scholar]
13.
Brühl CA, Gunsalam G, Linsenmair KE. Stratification of ants (Hymenoptera, Formicidae) in a primary rain forest in Sabah. Borneo.
J. Trop. Ecol. 1998,
14, 285–297.
[Google Scholar]
14.
DeVries PJ, Alexander LG, Chacon IA, Fordyce JA. Similarity and difference among rainforest fruit-feeding Butterfly communities in Central and South America.
J. Anim. Ecol. 2012,
81, 472–482. doi:10.1111/j.1365-2656.2011.01922.x.
[Google Scholar]
15.
Mena S, Kozak KM, Cárdenas RE, Checa MF. Forest stratification shapes allometry and flight morphology of tropical butterflies.
Proc. R. Soc. B 2020,
287, 20201071. doi:10.1098/rspb.2020.1071.
[Google Scholar]
16.
Basset Y, Aberlenc HP, Barrios H, Curletti G, Béranger JM, Vesco JP, et al. Stratification and diel activity of arthropods in a lowland rain forest in Gabon.
Biol. J. Linn. Soc. 2001,
72, 585–607.
[Google Scholar]
17.
Frenne PD, Zellweger F, Rodríguez-Sánchez F, Scheffers BR, Hylander K, Luoto M, et al. Global buffering of temperatures under forest canopies.
Nat. Ecol. Evol. 2019,
3, 744–749. doi:10.1038/s41559-019-0842-1.
[Google Scholar]
18.
Hallé F. Distribution verticale des métabolites secondaires en forêt équatoriale—une hypothèse. In Biologie d’une Canopée de Forêt Equatoriale—III. Rapport de la Mission d’ Exploration Scientifique de la Canopée de Guyane, Octobre—Décembre 1996; Hallé F, Ed.; Pro-Natura International & Opération Canopée: Paris, France, 1998; pp. 129– 138.
19.
Szarzynski J, Anhuf D. Micrometerological conditions and canopy energy exchanges of a neotropical rain forest (Surumoni–Crane Project, Venezuela).
Plant Ecol. 2001,
153, 231–239.
[Google Scholar]
20.
Cunningham-Minnick MJ, Roberts HP, Milam J, King DI. Sampling the understory, midstory, and canopy is necessary to fully characterize native bee communities of temperate forests and their dynamic environmental relationships.
Front. Ecol. Evol. 2024,
12, 1352266. doi:10.3389/fevo.2024.1352266.
[Google Scholar]
21.
Leksono AS, Nakagoshi N. A review of Vertical and Spatial Variations of Canopy Insects. In Secondary Forests in Central Japan and East Java, Indonesia, Proceeding of 6th ICGRC International Conference on Global Resource Conservation (ICGRC), Malang, Indonesia, 30 November 2015; ICGRC: Malang, Indonesia, 2015; pp. 21–25.
22.
Normann C, Tscharntke T, Scherber C. Interacting effects of forest stratum, edge and tree diversity on beetles.
For. Ecol. Manage. 2016,
361, 421–431. doi:10.1016/j.foreco.2015.11.002.
[Google Scholar]
23.
Weiss M, Didham RK, Procházka J, Schlaghamerský J, Basset Y, Odegaard F, et al. Saproxylic beetles in tropical and temperate forests–A standardized comparison of vertical stratification patterns.
For. Ecol. Manag. 2019,
444, 50–58. doi:10.1016/j.foreco.2019.04.021.
[Google Scholar]
24.
Ulyshen MD, Hanula JL. A Comparison of the Beetle (Coleoptera) Fauna Captured at Two Heights Above the Ground in a North American Temperate Deciduous Forest.
Am. Midl. Nat. 2007,
158, 260–278.
[Google Scholar]
25.
Hardersen S, Curletti G, Leseigneur L, Platia G, Liberti G, Leo P, et al. Spatio-temporal analysis of beetles from the canopy and ground layer in an Italian lowland forest.
Bull. Insectology 2014,
67, 87–97.
[Google Scholar]
26.
Lawrence E. Henderson’s Dictionary of Biology; Pearson Benjamin Cummings: Harlow, UK, 2008.
27.
King DA, Wright SJ, Connell JH. The contribution of interspecific variation in maximum tree height to tropical and temperate diversity.
J. Trop. Ecol. 2006,
221, 11e24.
[Google Scholar]
28.
Yamakura T. An empirical approach to the analysis of forest stratification. I. Proposed graphical method derived by using an empirical distribution function.
Bot. Mag. Tokyo 1987,
100, 109e128. doi:10.1007/BF02488317.
[Google Scholar]
29.
LaFrankie JV, Ashton PS, Chuyong GB, Co L, Condit R, Davies SJ, et al. Contrasting structure and composition of the understory in species-rich tropical rain forests.
Ecology 2006,
87, 2298–2305.
[Google Scholar]
30.
Draper FC, Costa FRC, Arellano G, Phillips OL, Duque A, Macía MJ, et al. Amazon tree dominance across forest strata.
Nat. Ecol. Evol. 2020,
5, 757–767.
[Google Scholar]
31.
Laurans M, Hérault B, Vieilledent G, Vincent G. Vertical stratification reduces competition for light in dense tropical forests.
Ecol. Manag. 2014,
329, 79–88.
[Google Scholar]
32.
Wesenberg J. Blühphänologie im Kronenraum eines tropischen Tieflandregenwaldes am Oberen Orinoco, Amazonas, Venezuela. Doctoral Dissertation, Fakultät für Biowissenschaften, Pharmazie und Psychologie, Universität Leipzig, Leipzig, Germany, 2004.
33.
Prance GT. Notes on the vegetation of Amazonia III. The terminology of Amazonian forest types subject to inundation.
Brittonia 1979,
31, 26–38.
[Google Scholar]
34.
Grimbacher PS, Stork NE. Vertical stratification of feeding guilds and body size in beetle assemblages from an Australian tropical rainforest.
Austral Ecol. 2007,
32, 77–85. doi:10.1111/j.1442-9993.2007.01735.x.
[Google Scholar]
35.
Sobek S, Tscharntke T, Scherber C, Schiele S, Steffan-Dewenter I. Canopy
vs. understory: Does tree diversity affect bee and wasp communities and their natural enemies across forest strata?
For. Ecol. Manag. 2009,
258, 609–615. doi:10.1016/j.foreco.2009.04.026.
[Google Scholar]
36.
Berkov A, Tavakilian G. Host utilization of the Brazil nut family (Lecythidaceae) by sympatric woodboring species of
Palame (Coleoptera, Cerambycidae, Lamiinae, Acanthocinini).
Biol. J. Linn. Soc. 1999,
67, 181–198.
[Google Scholar]
37.
Floren A. Sampling arthropods from the canopy by insecticidal knockdown. In Manual on Field Recording Techniques and Protocols for All Taxa Biodiversity Inventories, Part 1, Vol. 8; Eymann J, Degref J, Häuser C, Monje JC, Samyn Y, VandenSpiegel D, Eds.; ABC Taxa: Bruselles, Belgium, 2010; pp. 158–172.
38.
Watanabe M, Nagamitsu T, Itioka T, Meleng P, Kojima H. Species Diversity and Vertical Distributions of Weevils (Coleoptera, Curculionidae) Collected by Collision Traps with Floral-fragrance Attractants in a Bornean Rainforest.
Elytra 2017,
7, 491–498.
[Google Scholar]
39.
Medianero E, Valderrama A, Barrios H. Diversidad de insectos minadores y formadores de agallas en el dosel y sotobosque del bosque tropical.
Acta Zool. Mex. 2003,
1, 153–168.
[Google Scholar]
40.
Barrios H. Insect herbivores feeding on conspecific seedlings and trees. In Arthropods of Tropical Forests. Spatio-Temporal Dynamics and Resource Use in the Canopy; Basset Y, Novotny V, Miller SE, Kitching RL, Eds.; Cambridge University Press: Cambridge, UK, 2003; pp. 282–290.
41.
Nagamitsu T, Momose K, Inoue T, Roubik DW. Preference in flower visits and partitioning in pollen diets of stingless bees in an Asian tropical rain forest.
Res. Popul. Ecol. 1999,
41, 195–202. doi:10.1007/s101440050023.
[Google Scholar]
42.
Ramalho M. Stingless bees and mass flowering trees in the canopy of Atlantic forest: a tight relationship.
Acta Bot. Bras. 2004,
18, 37–47. doi:10.1590/s0102-33062004000100005.
[Google Scholar]
43.
Ulyshen MD, Soon V, Hanula JL. On the vertical distribution of bees in a temperate deciduous forest.
Insect Conserv. Diver. 2010,
3, 222–228. doi:10.1111/j.1752-4598.2010.00092.x.
[Google Scholar]
44.
Oliveira BF, Scheffers BR. Vertical stratification influences global patterns of biodiversity.
Ecography 2019,
42, 249–249. doi:10.1111/ecog.03636.
[Google Scholar]
45.
Schleuning M, Blüthgen N, Flörchinger M, Braun J, Schaefer H, Böhning-Gaese K. Specialization and interaction strength in a tropical plant-frugivore network differ among forest strata.
Ecology 2011,
92, 26–36.
[Google Scholar]
46.
Kirmse S. Diel activity patterns of a canopy-inhibiting beetle community (Coleoptera) in a Neotropical rainforest.
Fron. For. Glob. Chang. 2024,
7, 1370044. doi:10.3389/ffgc.2024.1370044.
[Google Scholar]
47.
Anhuf D, Motzer T, Rollenbeck R, Schröder B, Scarzynski J. Water budget of the Surumoni crane site (Venezuela).
Selbyana 1999,
20, 179–185.
[Google Scholar]
48.
Anhuf D, Winkler H. Geographical and ecological settings of the Surumoni-Crane-Project 784 (Upper Orinoco, Estado Amazonas, Venezuela).
Anz. Akad. Wiss. Wien, Math. Naturwiss. Kl. I 1999,
135, 3–23.
[Google Scholar]
49.
Morrone JJ. Monograph biogeographical regionalisation of the Neotropical region.
Zootaxa 2014,
3782, 1–110. doi:10.11646/zootaxa.3782.1.1.
[Google Scholar]
50.
Kirmse S. Beetle diversity (Coleoptera) in the canopy of a tropical lowland rainforest in Venezuela.
PeerJ 2024, accepted.
[Google Scholar]
51.
Lamarre GPA, Molto Q, Fine PVA, Baraloto C. A comparison of two common flight interception traps to survey tropical arthropods.
ZooKeys 2012,
216, 43–55.
[Google Scholar]
52.
Riley KNP, Browne RA, Erwin TL. Carabid beetle (Coleoptera, Carabidae) richness, diversity, and community structure in the understory of temporarily flooded and non-flooded Amazonian forests of Ecuador.
ZooKeys 2021,
1044, 831–876. doi:10.3897/zookeys.1044.62340.
[Google Scholar]
53.
Russo L, Stehouwer R, Heberling JM, Shea K. The composite insect trap: an innovative combination trap for biologically diverse sampling.
PLoS ONE 2011,
6, e21079. doi:10.1371/journal.pone.0021079.
[Google Scholar]
54.
Arndt E, Kirmse S. Seed-feeding ground beetle communities (Coleoptera: Carabidae: Harpalini) on fruit falls of
Miconia and
Goupia in South Venezuela.
Stud. Neotrop. Fauna Environ. 2002,
37, 151–157.
[Google Scholar]
55.
Paarmann W, Adis J, Stork N, Gutzmann B, Stumpe P, Staritz B, et al. The structure of ground beetle assemblages (Coleoptera: Carabidae) at fig fruit falls (Moraceae) in a terra firme rain forest near Manaus (Brazil).
Ecotropica 2001,
17, 549–561.
[Google Scholar]
56.
Hoekman D, LeVan KE, Ball GE, Browne RA, Davidson RL, Erwin TL, et al. Design for ground beetle abundance and diversity sampling within the National Ecological Observatory Network.
Ecosphere 2017,
8, e01744. doi:10.1002/ecs2.1744.
[Google Scholar]
57.
Jarosík V. Pitfall trapping and species abundance relationships: a value for carabid beetles (Coleoptera: Carabidae).
Acta Entomol. Bohemoslov. 1992,
89, 1–12.
[Google Scholar]
58.
Mommertz S, Schauer C, Kösters N, Lang A, Filser J. A comparison of D-vacsuction, fenced and unfenced pitfall trap sampling of epigeal arthropods in agroecosystems.
Ann. Zool. Fennici 1996,
33, 117–124.
[Google Scholar]
59.
Kirmse S. Structure and composition of a canopy-beetle community (Coleoptera) in a Neotropical lowland rainforest in southern Venezuela.
R. Soc. Open Sci. 2024,
11, 240478. doi:10.1098/rsos.240478.
[Google Scholar]
60.
Reichardt H. A synopsis of the genera of neotropical Carabidae (Insecta: Coleoptera).
Quaest. Entomol. 1977,
13, 346–493.
[Google Scholar]
61.
Bouchard P, Bousquet Y, Davies AE, Alonso-Zarazaga MA, Lawrence JF, Lyal CHC, et al. Family-group names in Coleoptera (Insecta).
ZooKeys 2011,
88, 1–972.
[Google Scholar]
62.
Kirmse S. Beetle diversity on 23 canopy-tree species in the Amazonas (dataset).
Dryad Digit. Repos. 2024. doi:10.5061/dryad.kprr4xhc7.
[Google Scholar]
63.
Hammer Ø, Harper DAT, Ryan PD. PAST: Palaeontological statistics software package for education and data analysis.
Palaeontol. Electron. 2001,
4, 9.
[Google Scholar]
64.
Hammond PM, Stork NE, Brendell MJD. Treecrown beetles in context: A comparison of canopy and other ecotone assemblages in a lowland tropical forest in Sulawesi. In Canopy Arthropods; Stork NE, Adis J, Didham RK, Eds.; Chapman & Hall: London, UK, 1997; pp. 184–223.
65.
Davis AJ, Sutton SL, Brendell MJD. Vertical distribution of beetles in a tropical rainforest in Sulawesi: The role of the canopy in contributing to biodiversity.
Sepilok Bull. 2011,
13, 59–83.
[Google Scholar]
66.
McCaig T, Sam L, Nakamura A, Stork NE. Is insect vertical distribution in rainforests better explained by distance from the canopy top or distance from the ground?
Biodiv. Conserv. 2019,
29, 1081–1103. doi:10.1007/s10531-019-01927-0.
[Google Scholar]
67.
Kirmse S, Chaboo CS. Flowers are essential to maintain high beetle diversity (Coleoptera) in a Neotropical rainforest canopy.
J. Nat. Hist. 2020,
54, 1661–1696. doi:10.1080/00222933.2020.1811414.
[Google Scholar]
68.
Wolda H, OBrien CW, Stockwell HP. Weevil Diversity and Seasonality in Tropical Panama as Deduced from Light-Trap Catches (Coleoptera: Curculionoidea).
Smithson. Contrib. Zool. 1998,
590, 79.
[Google Scholar]
69.
Charles E, Basset Y. Vertical stratification of leaf beetle assemblages (Coleoptera: Chrysomelidae) in two forest types in Panama.
J. Trop. Ecol. 2005,
21, 329–336.
[Google Scholar]
70.
Arndt E, Beutel RG, Will KW. 7.8. Carabidae Latreille, 1802. In Handbook of Zoology, Vol. IV Arthropoda: Insecta. Part 38: Coleoptera, Vol. 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga (Partim); Kristensen NP, Beutel RG, Leschen RAB, Eds.; Walter De Gruyter: Berlin, Germany; New York, NY, USA, 2005; pp. 119–146.
71.
Bellamy CL, Nelson GH. Chapter 41. Buprestidae (Leach, 1815). In American Beetles: Polyphaga: Scarabaeoidea through Curculionidea; Bd. 2; Arnett RH, Thomas RMC, Skelly PE, Frank JL, Eds.; CRC Press LLC: Boca Raton, FL, USA, 2002; pp. 98–112.
72.
Bezark LG, Monné MA. Checklist of the Oxypeltidae, Vesperidae, Disteniidae and Cerambycidae, (Coleoptera) of the Western Hemisphere. 2013. Available online: https://apps2.cdfa.ca.gov/publicApps/plant/bycidDB/wdocuments.asp?w=n (accessed on 30 August 2024).
73.
Liljeblad E. Monograph of the Family Mordellidae (Coleoptera) of North America, North of Mexico; University of Michigan Press: Ann Arbor, MI, USA, 1945.
74.
Pokon R, Novotny V, Samuelson GA. Host specialization and species richness of root-feeding chrysomelid larvae (Chrysomelidae, Coleoptera) in a New Guinea rain forest.
J. Trop. Ecol. 2005,
21, 595–604.
[Google Scholar]
75.
Scholz CH, Chown SL. The evolution of habitat use and diet in Scarabaeoidea: A phylogenetic approach. In Biology, Phylogeny, and Classification of Coleoptera. Papers Celebrating the 80th Birthday of Roy A. Crowson; Pakaluk J, Ślipiński SA, Eds.; Muzeum i Instytut Zoologii PAN: Warszawa, Poland, 1995; pp. 355–374.
76.
Johnson PJ. Elateridae. In American Beetles: Polyphaga: Scarabaeoidea through Curculionidea; Arnett RH, Jr., Thomas MC, Skelley PE, Frank JH, Eds.; CRC Press LLC: Boca Raton, FL, USA, 2002; Volume 2, pp. 160–173.
77.
Jolivet PHAH, Cox ML, Petitpierre E. Novel Aspects of the Biology of Chrysomelidae; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994.
78.
Kirmse S, Chaboo CS. Polyphagy and florivory prevail in a leaf-beetle community (Coleoptera: Chrysomelidae) inhabiting the canopy of a tropical lowland rainforest in southern Venezuela.
J. Nat. Hist. 2018,
52, 2677–2721. doi:10.1080/00222933.2018.1548666.
[Google Scholar]
79.
Erber D. Biology of Camptostomata Clytrinae–Cryptocephalinae–Chlamisinae–Lamprostomatinae. In Biology of Chrysomelidae; Jolivet PA, Petitpierre E, Hsiao TH, Eds.; Kluwer Academic: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 1988; pp. 513–552.
80.
Jolivet P. Food habits and food selection of Chrysomelidae. Bionomic and evolutionary perspectives. In Biology of Chrysomelidae; Jolivet PA, Petitpierre E, Hsiao TH, Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 1–24.
81.
Anderson RS. Curculionidae. In American Beetles. Polyphaga: Scarabaeoidea through Curculionoidea; Arnett RH, Jr., Thomas MC, Skelley P, Eds.; CRC Press LLC: Boca Raton, FL, USA, 2002; pp. 722–815.
82.
Berkov A. Chapter 16. Seasonality and Stratification: Neotropical Saproxylic Beetles Respond to a Heat and Moisture Continuum with Conservatism and Plasticity.
Saproxylic Insects:Divers. Ecol. Conserv. 2018, 547–578. doi:10.1007/978-3-319-75937-1_16.
[Google Scholar]
83.
Erwin TL. Forest Canopies, Animal Diversity. In Encyclopedia of Biodiversity; Levin SA, Ed.; Academic Press: Waltham, MA, USA, 2013; pp. 511–515.