1. Introduction
The respiratory tract is densely innervated by the nervous system, which allows it to respond flexibly to various environmental stimuli, physiological (respiration, coughing) and pathological processes (hypoxia, inflammatory responses) [
1,
2,
3,
4]. The nucleus of the solitary tract (NTS) is a key nucleus in the brainstem and is the main hub for perceiving and integrating visceral sensory information (). It receives afferent signals from the sensory neurons of the vagal nodose ganglion and the glossopharyngeal nerves and plays a key role in regulating respiratory, cardiovascular, and gastrointestinal function [
5]. Although the NTS is considered the central hub for sensing pulmonary allergens and pathogens, integrating this information and delivering it to other nuclei, its specific neural circuitry remains unclear. The neurocircuitry that regulates pulmonary diseases plays a key role in controlling airway hyperresponsiveness (AHR) and inflammatory responses. This regulation occurs through the interaction of the parasympathetic and sympathetic nervous systems [
6]. Airway constriction, or AHR, is known to be induced by inhaled irritants and allergens [
7]. However, it is still unknown whether NTS is associated with allergen−induced AHR.
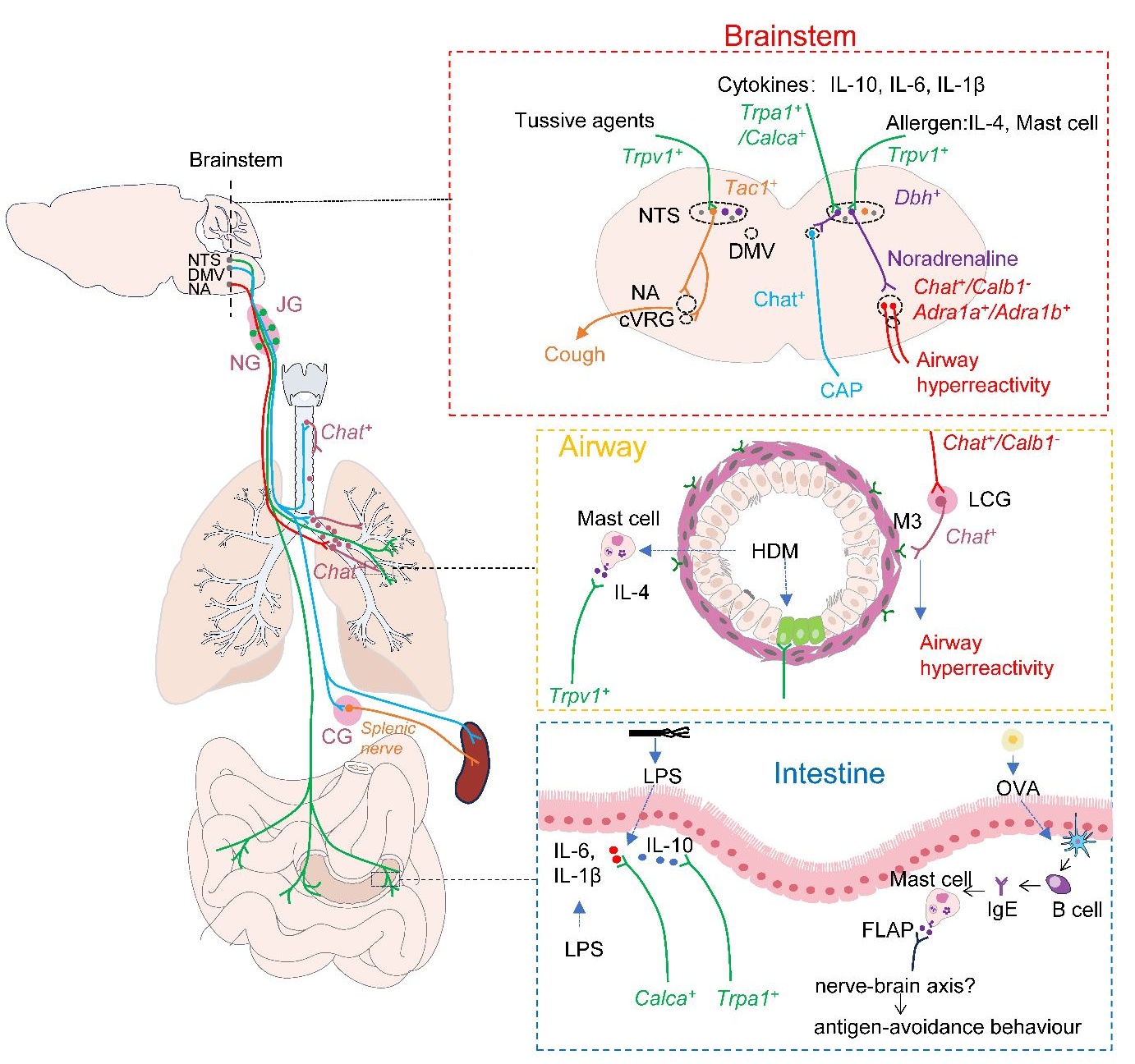
. The vagal interoceptive circuitry connecting peripheral organs to the brainstem mediates the body’s response to external stimuli. <i>Dbh</i><sup>+</sup> neurons in the NTS can receive signals from vagal <i>Trpv1</i><sup>+</sup> sensory neurons that sense lung allergens, such as IL−4 produced by HDM−induced mast cells. After integrating these signals, NTS <i>Dbh</i><sup>+</sup> neurons transmit them to <i>Chat</i><sup>+</sup> neurons in the NA via noradrenaline. The activated NA <i>Chat</i><sup>+</sup> neurons drive airway hyperreactivity through their interaction with pulmonary cholinergic ganglia. Additionally, NTS <i>Dbh</i><sup>+</sup> neurons can also receive signals from vagal <i>Trpa1</i><sup>+</sup> and <i>Calca</i><sup>+</sup> sensory neurons, which sense the anti−inflammatory factor IL−10 and the pro−inflammatory factors IL−6 and IL−1β, respectively. After integrating these signals, NTS <i>Dbh</i><sup>+</sup> neurons can suppress the production of pro−inflammatory factors in the spleen through traditional CAPs or direct projections from the DMV to the spleen. Besides <i>Dbh</i><sup>+</sup> neurons, NTS <i>Tac1</i><sup>+</sup> neurons can also receive signals from vagal <i>Trpv1</i><sup>+</sup> sensory neurons that sense tussive agents. After integration in NTS <i>Tac1</i><sup>+</sup> neurons, these signals are relayed to NA and caudal ventral respiratory group (cVRG) neurons, triggering the cough reflex. Moreover, mast cells are involved not only in sensing airborne allergens but also in sensing food allergens such as OVA. They transmit signals to the brain via the neuron−immune axis, eliciting antigen−avoidance behavior, although the specific neurons and central circuits involved in this recognition remain unclear. NTS: solitary tractus nucleus; DMV: dorsal motor nucleus of the vagus nerve; NA: nucleus ambiguus; cVRG: caudal ventral respiratory group; JG: jugular ganglion; NG: nodose ganglion; LCG: lung cholinergic ganglion; CG: celiac ganglion; CAP: cholinergic anti−inflammatory pathway; Dbh: dopamine beta−hydroxylase; Chat: Choline O−Acetyltransferase; Tac1: Tachykinin Precursor 1; Trpv1: transient receptor potential cation channel subfamily v member 1; Trpa1:transient receptor potential cation channel subfamily a member 1; Calca: calcitonin related polypeptide alpha; Adra1a/Adra1b: adrenoceptor alpha 1a/1b; Calb1: Calbindin 1; FLAP: 5−LO−activating protein; HDM: house dust mites; OVA: Ovalbumin.
In a recent issue of the journal Nature, Su et al. [
8] reported a complete allergen recognition and response circuit from the lungs to the brainstem and back to the lungs. They highlighted that the specific clusters of
Dbh+ neurons in NTS could integrate and transmit allergic signals. Specifically, in response to the allergen challenge, vagal
Trpv1+ neurons sense IL−4 produced by mast cells and relay the received signals to
Dbh+ neurons in the NTS. The activated
Dbh+ neurons release norepinephrine, which interacts with the noradrenergic receptors on
Chat+ neurons in the nucleus ambiguus (NA) to initiate the airway hyperresponsiveness reflex. These findings elucidate the role of vagal
Trpv1+ sensory neurons in allergen recognition and how the respiratory nervous centers (NTS and NA) control allergen−induced AHR, which may provide new intervention strategies for asthma treatment.
2. Dbh+ NTS Neurons Are Activated during HDM−Induced Asthma
To investigate whether an allergen (HDM) challenge in the lungs could trigger brainstem neuronal activation, Su et al. performed immunostaining for the immediate early factor FOS, a marker of neuronal activation. They found that the neurons in the NTS were activated only after repeated exposure to inhaled HDM (for 4 times) [
8]. To verify how NTS neurons sense repeated allergen exposure, Su et al. used mast cell−deficient
c−Kitw−sh/w−sh mice, anti−IL−4 neutralizing antibody administration, and unilateral vagotomy [
8]. These methods validate the involvement of mast cells, IL−4, and vagus nerve in activating NTS neurons. Recent studies have shown that, unlike chronic diseases such as asthma, NTS neurons respond very quickly to acute conditions such as sepsis, and sensory neurons in the NTS can respond in less than 2 h. This rapid response stems from the recognition of LPS−induced anti−inflammatory factor IL−10 and pro−inflammatory factors IL−6 and IL−1β by
Trpa1+ and
Calca+ vagal sensory neurons [
9]. Activated NTS
Dbh+ neurons relay messages to the dorsal motor nucleus of the vagus (DMV), which could inhibit splenic TNF−α production induced by LPS challenge via vagal celiac ganglion (CG) and their efferents (named cholinergic anti−inflammatory pathway, CAP) [
10]. More importantly, a recent study has confirmed the direct projection from DMV to the spleen [
11].
The NTS contains various types of neurons, each potentially serving distinct functions. The researchers in this study also used single−nucleus transcriptome analysis to confirm the gene expression profiles of activated NTS neurons during asthma. They found that
Dbh+ neurons are preferentially activated to mediate AHR [
8]. In another recent study, researchers found that
Tac1+ neurons in the NTS are a key component of the airway−vagal−brain neural circuit, which controls cough−like defense behavior in mice [
12], suggesting that the NTS
Tac1+ neurons recognize cough−inducing stimuli. Therefore, we can hypothesize that different types of neurons in NTS are involved in recognizing different substances and performing different functions. The same type of neuron can perceive different substances, effectively expanding the body’s ability to respond to complex and variable external stimuli.
3. Dbh+ NTS Neurons Specifically Trigger Airway Hyperreactivity
Subsequently, the researchers modulated
Dbh+ NTS neurons through chemical ablation, genetic ablation, and chemogenetic inactivation/activation. The results unequivocally demonstrated that
Dbh+ NTS neurons play a crucial role in allergen−induced airway hyperreactivity. Still, they are not involved in regulating HDM−induced goblet cell metaplasia, immune cell infiltration, or the expression levels of
Il4,
Il5, and
Il13. Additionally, modulating
Dbh+ NTS neurons did not cause any significant changes in respiratory parameters such as minute ventilation, respiratory frequency, tidal volume, or metabolic rate. These findings suggest that when allergens activate
Dbh+ NTS neurons in the brainstem, they trigger airway smooth muscle contraction through a specific neural circuit, leading to airway hyperresponsiveness (AHR).
To identify first−order sensory neurons, Su et al. injected TRAP2 (targeted recombination in active populations) mice with rabies virus into NTS. They found that vagal
Trpv1+ sensory neurons could recognize repeated exposure to allergens. Thus, vagal
Trpv1+ neurons detect IL−4 produced by mast cells and mediate allergen−induced AHR [
8]. Similarly, in the intestine, 5−LO−activating protein (FLAP) released by mast cells is also involved in sensing intestinal allergens and mediating antigen avoidance behaviors [
13]. The findings support that the activation of
Dbh+ neurons in NTS is primarily driven by inflammatory cytokines produced by immune cells after
Trpv1+ vagal sensory neurons sensing allergens.
Tamari et al. reported that β−CGRP
+ vagal sensory neurons could suppress group 2 innate lymphoid cell (ILC2) function and allergic airway inflammation via the JAK1−β−CGRP signaling pathway [
14]. In light of these findings with the results in the Nature paper, specifically that the regulation of
Dbh+ NTS neurons in the brainstem does not alter type 2 immune responses, we speculate that various types of sensory neurons within the vagus ganglion may receive distinct signals and relay them to the central nervous system via different neural circuits. Following central integration, these signals are relayed to diverse effector cells, such as airway smooth muscle cells, ILC2s, or other cell types, ultimately resulting in differential physiological changes. The above findings suggest that further research is warranted to investigate projections and functions of the nuclei in the central nervous system and peripheral sensory neurons in the pathogenesis of lung disease.
It has been reported that the downstream targets of the NTS include the DMV and the NA, which are the origins of vagal motor fibers. These fibers regulate visceral functions by directly acting on target cells or projecting to postganglionic neurons in target organs [
15]. Following this idea, the researchers used viral tracing technology. They found that
Dbh+ neurons in the NTS project to the NA, not the DMV, and relay allergen signals to postganglionic neurons that directly drive airway constriction through norepinephrine (NE) released by
Dbh+ neurons interacting with NE receptors in the NA. Notably, the neurons in NA that regulate HDM−induced AHR are
Chat+Calb1(Calbindin)
− neurons. One study showed that NA
Calb1+ neurons project to pulmonary ganglia and mediate the diving reflex, which involves the simultaneous presence of bradycardia and bronchoconstriction after water immersion [
16]. This indicates that different populations of NA neurons may innervate pulmonary ganglia, which specifically affect airway constriction and respiratory reflexes.
The authors used the Flexivent assay to assess airway hyperresponsiveness in this work. They administered intratracheal increasing doses of methacholine to anesthetized mice to induce airway constriction. We must note that exogenous methacholine may not be able to mimic the effects of endogenous acetylcholine released by parasympathetic nerves. How to demonstrate that parasympathetic-derived acetylcholine contributes to airway smooth muscle contraction warrants further investigation.
4. Summary and Future Outlook
The NTS can be activated by various diseases, including asthma, sepsis, cough, acute lung injury, and diabetes () [
5,
9,
12,
17]. The modulatory role of the lung−NTS
Dbh+ interoceptive circuit in AHR makes it a potential target for future asthma therapies. Still, its impact on other diseases also needs to be further studied. In addition to NTS
Dbh+ and
Tac1+ neurons, there are many other classified neurons in the NTS whose function must also be determined. In addition, there are unresolved issues, such as the differences and the commonalities between the acute response of NTS
Dbh+ neurons to LPS versus the chronic response of repeated allergen exposure−induced cytokines.
Furthermore, the function of cholinergic pulmonary ganglia in the lungs needs to be clarified. For example, while
Chat+Calb1− neurons in the NA regulate HDM−induced AHR by projecting to a small number of cholinergic pulmonary ganglia, the role of
Calb1+ neurons projecting to a greater proportion of cholinergic pulmonary ganglia in allergic diseases or other lung conditions remains to be explored. Understanding the lung-vagus-brainstem circuit, which plays a key role in regulating AHR, could open up new therapeutic opportunities for preventing and treating lung diseases.
Acknowledgments
We thank all members of the Su lab for their helpful comments and discussion.
Author Contributions
R.W. and J.C. wrote the original manuscript. J.C. prepared the figure, and X.S. reviewed and edited the manuscript.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This work was supported by NSFC programs (82241042 and 81970075); National Key Research and Development Program of China (No.2022YFC2304702); Science and Technology Commission of Shanghai Municipality (20DZ2261200); China Postdoctoral Science Foundation (2024M750522).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1.
Wang H, Yu M, Ochani M, Amella CA, Tanovic M, Susarla S, et al. Nicotinic acetylcholine receptor alpha7 subunit is an essential regulator of inflammation.
Nature 2003,
421, 384−388. doi: 10.1038/nature01339.
[Google Scholar]
2.
Zhao C, Chen J, Liu Z, Liang H, Chen X, Cheng L, et al. Activation of nicotinic acetylcholine receptor alpha7 subunit limits Zika viral infection via promoting autophagy and ferroptosis. Mol. Ther. 2024, 32. doi:10.1016/j.ymthe.2024.05.037.
3.
Naqvi KF, Mazzone SB, Shiloh MU. Infectious and Inflammatory Pathways to Cough.
Annu. Rev. Physiol. 2023,
85, 71−91. doi: 10.1146/annurev−physiol−031422−092315.
[Google Scholar]
4.
Wen J, Sun J, Tang Y, Zhao J, Su X. Cholinergic alpha7 nAChR signaling suppresses SARS−CoV−2 infection and inflammation in lung epithelial cells.
J. Mol. Cell Biol. 2024,
15, mjad048. doi:10.1093/jmcb/mjad048.
[Google Scholar]
5.
Chen J, Lai X, Song Y, Su X. Neuroimmune recognition and regulation in the respiratory system.
Eur. Respir.Rev. An. Off. J. Eur. Respir. Soc. 2024,
33, 240008. doi:10.1183/16000617.0008−2024.
[Google Scholar]
6.
Nassenstein C, Krasteva−Christ G, Renz H. New aspects of neuroinflammation and neuroimmune crosstalk in the airways.
J. Allergy Clin. Immunol. 2018,
142, 1415–1422. doi:10.1016/j.jaci.2018.09.011.
[Google Scholar]
7.
Bradding P, Porsbjerg C, Côté A, Dahlén SE, Hallstrand TS, Brightling CE. Airway hyperresponsiveness in asthma: The role of the epithelium.
J. Allergy Clin. Immunol. 2024,
153, 1181–1193. doi:10.1016/j.jaci.2024.02.011.
[Google Scholar]
8.
Su Y, Xu J, Zhu Z, Chin J, Xu L, Yu H, et al. Brainstem
Dbh+ neurons control allergen−induced airway hyperreactivity.
Nature 2024,
631, 601–609. doi:10.1038/s41586−024−07608−5.
[Google Scholar]
9.
Jin H, Li M, Jeong E, Castro−Martinez F, Zuker CS. A body−brain circuit that regulates body inflammatory responses.
Nature 2024,
630, 695–703. doi:10.1038/s41586−024−07469−y.
[Google Scholar]
10.
Tracey KJ. The inflammatory reflex.
Nature 2002,
420, 853–859. doi:10.1038/nature01321.
[Google Scholar]
11.
Zhu X, Huang JY, Dong WY, Tang HD, Xu S, Wu Q, et al. Somatosensory cortex and central amygdala regulate neuropathic pain−mediated peripheral immune response via vagal projections to the spleen.
Nat. Neurosci. 2024,
27, 471–483. doi:10.1038/s41593−023−01561−8.
[Google Scholar]
12.
Gannot N, Li X, Phillips CD, Ozel AB, Uchima Koecklin KH, Lloyd JP, et al. A vagal−brainstem interoceptive circuit for cough−like defensive behaviors in mice.
Nat. Neurosci. 2024,
27, 1734−1744. doi:10.1038/s41593−024−01712−5.
[Google Scholar]
13.
Plum T, Binzberger R, Thiele R, Shang F, Postrach D, Fung C, et al. Mast cells link immune sensing to antigen−avoidance behaviour.
Nature 2023,
620, 634–642. doi:10.1038/s41586−023−06188−0.
[Google Scholar]
14.
Tamari M, Del Bel KL, Ver Heul AM, Zamidar L, Orimo K, Hoshi M, et al. Sensory neurons promote immune homeostasis in the lung.
Cell 2024,
187, 44–61.e17. doi:10.1016/j.cell.2023.11.027.
[Google Scholar]
15.
Travagli RA, Anselmi L. Vagal neurocircuitry and its influence on gastric motility.
Nat. Rev. Gastroenterol. Hepatol. 2016,
13, 389–401. doi:10.1038/nrgastro.2016.76.
[Google Scholar]
16.
Veerakumar A, Yung AR, Liu Y, Krasnow MA. Molecularly defined circuits for cardiovascular and cardiopulmonary control.
Nature 2022,
606, 739–746. doi:10.1038/s41586−022−04760−8.
[Google Scholar]
17.
Muscogiuri G, DeFronzo RA, Gastaldelli A, Holst JJ. Glucagon−like Peptide−1 and the Central/Peripheral Nervous System: Crosstalk in Diabetes.
Trends Endocrinol. Metab. TEM 2017,
28, 88–103. doi:10.1016/j.tem.2016.10.001.
[Google Scholar]