1. Introduction
Following the World Health Organization report on the public health response to dementia [
1], it was estimated that in 2019 nearly 55 million people worldwide were living with dementia and by 2050 this number is projected to skyrocket to a staggering 139 million. This projection underscores the urgent need to address the impact of neurodegeneration and reduce its burden. One approach to preventing neurodegenerative diseases is to try to understand their onset and do an early diagnosis. It is critical to concentrate on preventative interventions that might possibly postpone or even prevent the beginning of neurodegenerative illnesses. Early detection is essential in this context since it allows for prompt intervention and the adoption of relevant lifestyle changes or medications. In that respect, studies conducted in recent years have generated a body of evidence indicating that classical MHC class I (MHC-I, HLA-I in humans, H-2 in mice) molecules are of paramount importance for correct brain development and homeostasis (see below). However, the molecular mechanisms used are far from being understood. One of the reasons for this riddle is likely related to the structural and highly polymorphic molecular characteristics of MHC-I molecules.
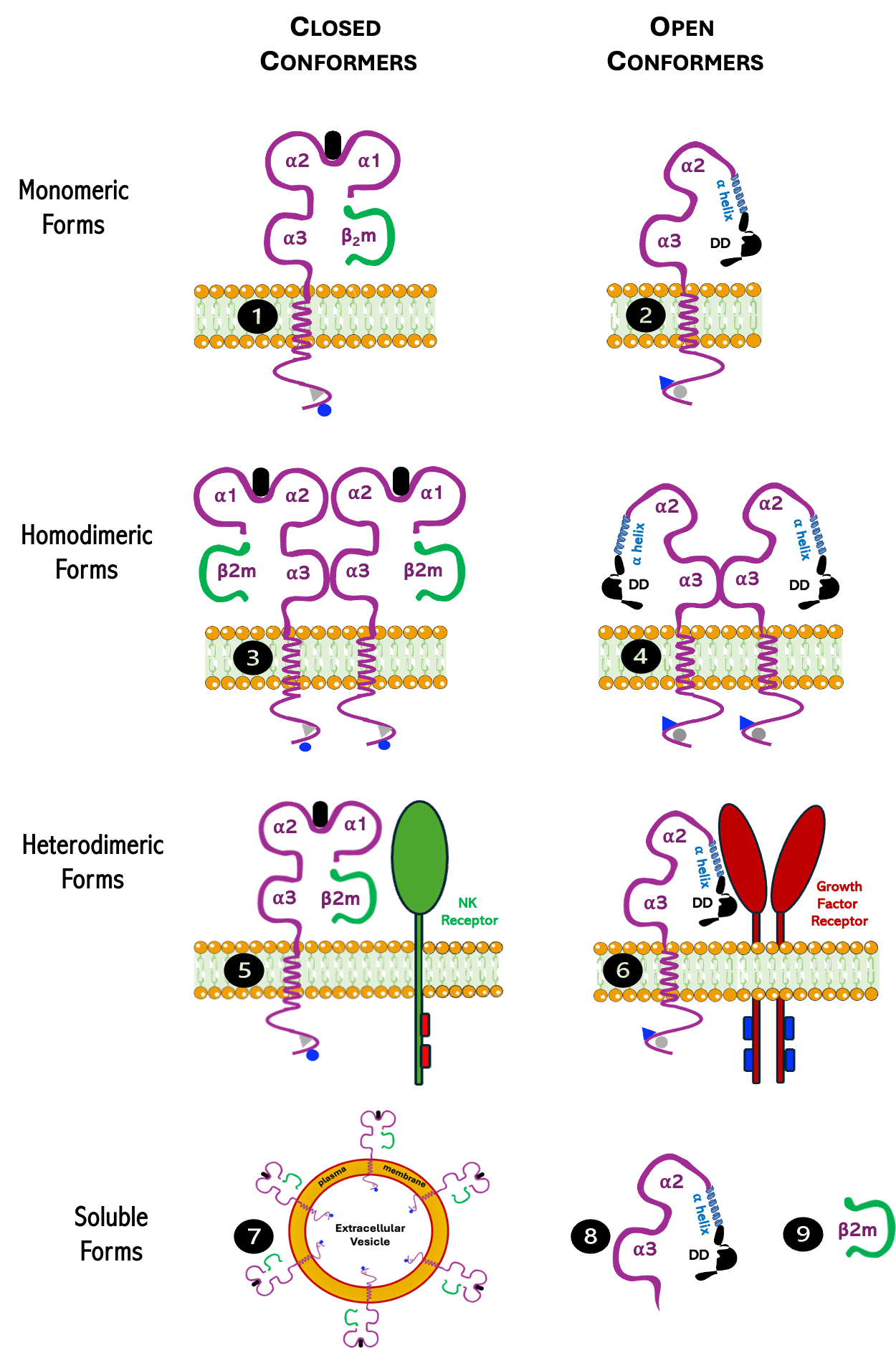
Briefly, classical MHC-I molecules are synthesized and assembled in the endoplasmic reticulum (ER) and then transported to the plasma membrane as closed conformers constituted by a heavy chain (αHC), a light chain (β2m) and an 8–11 amino acid peptide, where they can stay for about 6–7 h. During its lifespan they may eventually be recognized by the T cell receptor (TCR) of CD8+ T cells or by killer cell Ig-like receptors (KIR) and leukocyte Ig-like receptors (LILR) expressed by NK cells and effector-memory CD8+ T cells or be endocytosed and either recycled back to the plasma membrane or degraded [
2]. Expression of MHC-I molecules is regulated through the activation of the enhanceosome, a group of DNA regulatory sequences that includes enhancer A, IFN-stimulated response element, the SXY modules, and the Class I trans-activator protein, NLRC5 [
3]. This group of sequences is activated by extracellular signals, namely cytokines. The most important are IFN-γ and TNF-α, but other cytokines such as IFN-α, IFN-β and IL-4 also have some degree of influence [
4].
Noteworthy, the MHC-I molecules expressed at the cell surface of nucleated cells exist in a physiological equilibrium between β2m-associated (closed) and β2m-free (open) conformers. Conditions such as an increase in metabolic activity, cell growth and inflammation have been proved to be inducers of the open conformers and the release of β2m, and several studies have suggested that this equilibrium is influenced by nutritional and metabolic demands [
5,
6], and references herein. In this regard, it is important to mention that both the closed and the open conformers can interact in
cis (
i.e., interactions that take place in the plasma membrane of the same cell) with themselves, forming homodimers, or with other receptors, forming heterodimers, including receptors for growth factors and hormones (e.g., insulin, epidermal growth factor, fibroblast growth factor, glucagon, IL-15,
etc.), some of which are expressed by neurons and other CNS cells, and have important functional implications for the cell, reviewed in [
5,
6]. For instance, it is well known that the insulin receptor (IR) triggers intracellular signals involved in the regulation of synapse number, dendritic plasticity, and circuit function in vivo [
7]. Importantly,
cis-associations between the IR and, most likely, open MHC-I conformers fine-tune these intracellular signals in B-LCL cell lines by increasing the affinity of IR for insulin, enhancing phosphorylation of insulin receptor substrate-1 (IRS-1) and activating of phosphoinositide 3-kinase (PI3K) [
8]. In this regard, studies in mouse hippocampal neurons showed that MHC-I negatively regulates synapse density by inhibiting insulin receptor signaling [
9]. Even though the latter study suggests that MHC-I can influence insulin receptors in trans, it does not rule out the possibility that
cis-interactions can occur [
9], which deserves to be further dissected, especially taking into account the importance of this insulin-related sensitivity in the aging and diseased brain [
9]. Besides, both single closed and single open conformers and homodimers can interact in
cis and in
trans with a variety of inhibitory and activating NK receptors both in mice (e.g., Ly49 and PIR members) and humans (e.g., KIR, LILR, and CD94–NKG2 members), modulating important cellular functions [
10]. Of note, practically all mice H-2 class I alleles and human HLA-I alleles can establish these cis-interactions with a myriad of receptors, immune and non-immune [
11]. Moreover, the MHC-I molecules present in the plasma membrane, whether in a closed or an open conformation, can be released into the extracellular environment as “soluble” MHC-I molecules that can be found in a number of biological fluids, including plasma, saliva, cerebrospinal fluid (CSF), and others [
12]. In this respect, at the immunological level, a number of
in vitro studies have demonstrated that these soluble MHC-I molecules are immunosuppressor and tolerogenic factors capable of inhibiting NK and CD8+ T cell cytotoxic responses [
6]. Therefore, to shed light on the mechanisms used by MHC-I molecules to regulate essential biological processes in the brain, it is necessary to take into account each and every possible player just mentioned and the receptors and proteins they interact with ().
Here, we give a brief account of the studies that led to a non-immunological scenario where MHC-I molecules play essential roles in the regulation of brain homeostasis and that alterations in the expression of MHC-I molecules are associated with cognitive disorders and dementia. The major focus will be on neuroregeneration, synaptic plasticity, and signaling. This opinion article is based on the premise that the MHC-I molecules present at the plasma membrane of all nucleated cells, through their capacity to exist in a physiological equilibrium between closed and open conformers, are gatekeepers that fine-tune intracellular signals transmitted by the binding of nutrients and hormones to their receptors, and intracellular signals transmitted by inhibitory and activation receptors via
cis–
trans interactions [
5,
6]. Understanding when and how these polymorphic closed and open conformers influence brain homeostasis, healthy aging and the balance between neurodegeneration and neuroregeneration can open new avenues of research aiming at preserving cognitive function and ultimately to reduce its prevalence and the detrimental effect of neurodegenerative diseases such as Alzheimer’s and Parkinson’s diseases. Thus, MHC-I molecules, the conformations they can adopt, their molecular partners and the signaling pathways they modulate may become biomarkers of cognitive decline and, at the same time, putative targets for the development of innovative therapies. Based on the different forms that may exist during the lifetime of classical MHC-I molecules (), akin to the functional implications described so far, resulting from
cis–
trans interactions between the MHC-I forms and their identified ligands in the CNS, we will end by proposing some strategies for future studies.
. Model illustrating the nine different forms (❶ to ❾) that may exist during the life-time of a classical MHC class I molecule at the plasma membrane. Forms are listed as closed conformers (❶, ❸, and ❺), open conformers (❷, ❹ and ❻), and soluble forms (❼, ❽ and ❾). The extracellular domains of each form are indicated. Also indicated are the cytoplasmic conserved residues tyrosine 320 (Y320, triangle) and serine 335 (S335, circle). When Y320 or S335 are phosphorylated, are in blue. When they are dephosphorylated are in grey. Note that the transition closed to open conformer is associated with phosphorylation of Y320 and dephosphorylation of S335. The ordered ⍺ helix subdomain of the ⍺1 domain, and the disordered (DD) subdomain of the ⍺1 domain are indicted. Modified from ref. [<a href="#B6" class="html-bibr">6</a>].
2. A Role for Classical MHC Class I Molecules in Neuroregeneration
During recent years, a body of evidence has been accumulated indicating that classical MHC-I molecules are of paramount importance for correct brain development and homeostasis. Thanks to the seminal studies by the groups of Carla Shatz and Lisa Boulanger, it is presently known that MHC-I molecules are expressed by all brain parenchymal cells, including neurons [
13,
14]. These studies demonstrated that expression of MHC-I molecules was regulated by neuronal electrical activity and that their presence was important for synaptic plasticity and pruning during normal development. It was also found that expression of MHC-I molecules was increased by inflammatory cytokines, such as IFN-γ, and seizures. After these breakthrough findings, a myriad of studies using manipulated animal models followed.
Studies performed by the group of Staffan Cullheim examining the role of MHC-I molecules in neuroregeneration after nerve transection showed that mice lacking expression of MHC-I molecules exhibited more extensive removal of synapses and a significantly smaller population of motoneurons reinnervating the transected sciatic nerve than wild-type (WT) mice, suggesting that the absence of MHC-I molecules compromised the capacity of neurons to regenerate axons [
15]. A similar study using three different mice strains (
i.e., A/J, BALBc and B6) revealed enthralling results [
16]. After axotomy of motoneurons, A/J mice (Kk, Dd, Ld) displayed a robust increase of MHC-I, especially in the surroundings of lesioned motoneurons, revealed astroglial reaction, and faster synaptic elimination, resulting in a more effective regenerative outcome. The authors discussed that this transitory synaptic pruning would allow the cell to concentrate its resources and efforts on processes leading to structural restoration of lesioned axons. Similar results, although less marked, were observed in the BALBc mice (Kd, Dd, Ld). In marked contrast, B6 mice (Kb, Db, null for Ld) displayed a lower upregulation of MHC-I molecules in the spinal cord, a lower astroglial reaction, a slower synaptic pruning, and a poor regenerative potential [
16]. However, these results must be interpreted in the context of the time that these phenomena occur (e.g., acute, after the injury), as well as the specificity of the neurons in which this synaptic stripping occurs (e.g., inhibitory vs. excitatory neurons) [
15,
16]. This means that MHC-I needs to be carefully regulated for effective neuroregeneration. Noteworthy, the study of the three different mice strains unveiled three remarkable facts. First, the neuroregenerative process in mice seems to be influenced by the MHC-I haplotype. Second, upregulation of MHC-I molecules after the lesion positively influences neuroregeneration. Third, the absence of H-2Ld molecules in the B6 mice, which show poor regenerative potential, suggests that this allele may play an important role in the regenerative process. Interestingly, H-2Ld molecules have a tendency to lose β2m and turn into open conformers, which may result in the formation of homodimers or heterodimers [
17,
18,
19]. Thus, the formation of these new structures may have implications in the biological processes regulating neuroregeneration, perhaps by inducing signals that result in the secretion of neuroregenerative factors. In agreement with this line of thought, IFN-γ, which is a potent inducer of MHC-I expression and is upregulated in CNS after injury, appears to play a neuroprotective role. Accordingly, IFN-γ knock-out mice exhibited neuronal death and a reduced number of presynaptic terminals [
20].
In accord with these results, an elegant study by the group of Daniel Kaufman using mutant mice showed that increased expression of MHC-I molecules only on neurons was associated with significantly better recovery of locomotor abilities after spinal cord injury than WT mice, pointing again to MHC-I molecules as promoters and/or fine-tuners of neuroregeneration [
21]. In another study, using transgenic humanized mice expressing HLA-A11 or HLA-B27 molecules to ascertain their role in persistent CNS injury versus repair following CNS viral infection by a picornavirus [
22], it showed that unlike class I deficient mice, which died 18 to 21 days post-infection, both HLA-A11+ and HLA-B27+ transgenic mice significantly controlled virus expression by 45 days post-infection and did not develop late-onset spinal cord demyelination. Remarkably, while the HLA-A11+ transgenic mice showed persistent severe hippocampal and cortical injury, the HLA-B27+ transgenic mice showed almost complete repair of the brain. These results reinforce the notion that the neuroregenerative process is positively influenced by human MHC-I alleles, namely HLA-B27. Interestingly, the human HLA-B27 molecule, like the mouse H-2Ld, has a tendency to lose β2m and become open HLA-B27 conformers, which can form dimers capable to interacting with inhibitory receptors, including KIR and LILR [
6]. By analogy with the mice model, the better neuroregenerative process seen in humanized HLA-B27 mice could result from
cis and/or
trans interactions of the open HLA-B27 conformers with receptors involved in the neuroregenerative process. Additional evidence that upregulation of MHC-I molecules confers neuroprotection was shown by a recent study carried out in SOD1G93A transgenic mice, used as a model for amyotrophic lateral sclerosis, subjected to subcutaneous applications of IFN-β, a cytokine that like IFN-γ, upregulates MHC-I expression. The results showed that the IFN-β-mediated upregulation of MHC-I resulted in a significant increase in neuronal survival and synaptic preservation in the ALS presymptomatic period [
23]. Contrasting with these results, a recent study has shown that neuronal accumulation of ApoE triggers aberrant upregulation of MHC-I expression, which in turn drives tau pathology and the selective destruction of individual synapses and neurons and leads to selective neurodegeneration in Alzheimer’s disease (AD) [
24]. The apparent discrepancy between studies can be explained by the fact that the mouse model used in the later study was markedly different and expressed aberrantly high levels of MHC-I molecules. This brings about the important issue of the possible existence of a threshold of MHC-I expression above which MHC-I molecules are harmful and neurodegeneration rather than neuroregeneration prevails, as seen in other mouse models [
25]. In any case, this issue warrants further investigation to study if the levels of neuronal expression of MHC-I molecules could be modulated and accurately quantitated to determine which level of MHC-I expression favors neuroregeneration instead of neurodegeneration.
3. A Role for Open MHC-I Conformers in Synaptic Plasticity and Neuronal Growth
In a different set of studies, Kimberly McAllister’s group investigated the role of MHC-I molecules in the establishment of synaptic connections during development. Studies conducted in the rat visual cortex showed that MHC-I molecules are present both pre-synaptically and post-synaptically at all ages examined, suggesting that they have a role on both synaptic sides, perhaps regulating connectivity during and after the establishment of cortical connections [26]. In addition, by using rat cortical neuron cultures and mice neurons from the cortex of newborn B6 mice (Kb, Db, null for Ld) or mice lacking β2m, the same group unveiled important aspects of the biological role of MHC-I molecules on brain homeostasis [27]. Thus, both closed and open MHC-I conformers were expressed by glutamatergic neurons. In addition, the authors suggested that it was the pool of open MHC-I conformers, and not the pool of closed conformers, that was critical for the negative regulation of glutamatergic synapse density during cortical development in young, but not older cortex, likely through cis–trans interactions with receptors involved in synapse formation. In other words, the proportion of open conformers in relation to the closed ones MHC, dictated by the physiological equilibrium closed ⇌ open conformers [5,6], seems critical for MHC-I signaling and function in neurons. However, it is not clear from this study how the changes in open and closed conformers were quantitated, raising the possibility that there may be alternative interpretations to explain the phenomena observed. On the other hand, a recent study addressing the relationship between amyloid-β (Aβ), MHC-I expression and AD showed that the closed MHC-I conformers present on the cell surface of neuronal cells were dissociated after treatment with Aβ oligomers, leading to a decrease in closed MHC-I molecule expression, an effect also observed in brains of AD patients [28]. As expected, MHC-I disassembly led to the release of β2m and to the generation of open MHC-I conformers at the plasma membrane. From this study, it was concluded that the decrease in closed MHC-I conformers negatively impacted the signaling resulting from the interaction with NCAM1. This neuronal receptor is involved in the development of the nervous system by regulating neurogenesis, neurite outgrowth, and cell migration [29]. In line with these results, a recent study using a mice model of AD as well as human brain specimens showed that aggregates of β2m and Aβ are increased in the brains of AD patients and AD mice and trigger neurodegeneration, pointing to β2m as a neurodegenerative factor contributing to AD pathogenesis [30]. In that respect, it is of note that soluble β2m has been shown to act as an endogenous N-methyl-D-aspartate receptor (NMDAR) antagonist impairing synaptic function [31].
On the other hand, a series of interesting studies performed by Kaufman’s group showed that soluble MHC-I molecules were involved in the regulation of neurogenesis. Thus, by using recombinant soluble MHC-I molecules on cultured embryonic retinas, it was demonstrated that self-soluble MHC-I molecules (sMHC-I) induced greater neuroinhibitory signals than non-self sMHC-I, resulting in inhibition of neurite outgrowth [32]. Importantly, this study showed that inhibition of neurite outgrowth was mediated by closed MHC-I molecules regardless of the nature of the bound peptide (i.e., self vs. non-self). These observations strongly suggest that the inhibitory receptor(s) for the closed MHC-I conformers likely recognize conserved regions of the self-MHC-I molecules outside the peptide binding groove. Whether this recognition implies that neurons are “educated” to recognize self-MHC I, like it happens in the immunological system with NK cells, as initially proposed by Daniel Kaufman [32], is an intriguing question that deserves further investigation. This role was confirmed by using cultures of embryonic mouse retina within a short distance from thalamic explants obtained from WT mice or NSE-H-2Db transgenic mice, whose neurons express high levels of H-2Db molecules [33]. These studies showed that retinal neurites outgrew to form connections with WT but not with NSE-H-2Db thalamic explants due to growth inhibition. Notably, the inhibitory effect was shown to be mediated by soluble closed H-2Db conformers, regardless of the bound peptide, which reinforces the view put forward above that the inhibitory receptor(s) for the soluble closed MHC-I conformers likely recognize conserved regions of the self-MHC-I molecules outside the peptide binding groove. A third study by the same group concluded that a positive effect of MHC-I molecules on neuritogenesis might be mediated by cis-interactions between open H-2Db conformers and cell surface receptors (e.g., IR, etc.), reinforcing the important role of the open conformers on brain homeostasis [34]. For a more detailed description of the cis-interaction between MHC-I molecules and the IR see [6] (). In marked contrast, trans-interactions of closed H-2Db conformers with surface receptors caused inhibition of neurite growth, possibly by interacting with inhibitory surface receptors [34]. These data are enlightening because they showed for the first time a role for soluble closed MHC-I molecules in regulating neurite growth, as well as the importance of studying both closed and open MHC-I conformers. In this regard, it is noteworthy that our recent study in humans showed a high variability in the levels of soluble HLA-I molecules in the plasma of elderly people differing in their cognitive status. This study revealed, for the first time, that the coexistence of HLA-A23 or HLA-A24 alleles and dementia in elderly people was strongly associated with high levels of sHLA-I molecules [35]. Unlike the immunosuppressor activities of soluble MHC-I molecules described among cells of the immunological system (see above), the increased levels of soluble closed HLA-A23 or HLA-A24 may negatively contribute to neuroregeneration, and be considered potential biomarkers of neurodegeneration.
4. MHC-I Molecules Fine-Tune Signaling and Influence Brain Homeostasis
In a series of noteworthiest studies, the group of Lisa Boulanger provided the first clues to ascertain the mechanism(s) whereby MHC-I molecules influence brain homeostasis. By using WT and MHC-I deficient B6 mice (Kb, Db, null for Ld), the authors showed that endogenous MHC-I molecules tonically inhibited NMDAR function and controlled α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) trafficking in the mammalian CNS [
36]. This finding is of paramount importance for understanding the role of MHC-I molecules in brain homeostasis because NMDA receptors play an integral role in synaptic plasticity and are involved in a process called excitotoxicity, which can be harmful to neurons. On the other hand, AMPA receptors mediate fast excitatory synaptic transmission and are critical for normal brain function. This study strongly suggests that endogenous MHC-I expression could function as a neuroprotective factor within a certain level of expression, as mentioned above. In an ensuing study by the same group, it was shown that normal MHC-I expression is essential for NMDAR-dependent hippocampal synaptic depression and hippocampus-dependent memory, suggesting that MHC-I molecules are essential for normal cognition [
37]. In agreement with this view, Boulanger’s group also revealed a new and potential mechanism by which MHC-I can regulate neuronal connectivity by showing that endogenous MHC-I molecules regulate hippocampal synapse density by fine-tuning neuronal IR signaling after unmasking a cryptic epitope in the cytoplasmic tail of the IR [
9]. The authors raised the possibility that the action of MHC-I molecules could be mediated by soluble forms of MHC-I secreted for the neuronal cell surface, which is in agreement with the studies of Kaufman’s group, but the possibility that the interaction is in
cis with cell surface open MHC-I conformers cannot be ruled out. Further studies by Boulanger’s group identified cryptic protein interaction motifs in the cytoplasmic domain of MHC-I molecules, encompassing conserved tyrosine and serine residues, which could take part in protein-protein interactions at neuronal synapses [
38]. More recently, two recent reports from the group of Wilfred Jefferies and Dara Dickstein showed the importance of the cytoplasmatic tail of MHC-I molecules in synaptic signaling and its impact on the modulation of synaptic AMPA-type glutamate receptors (AMPARs). Thus, by replacing cytoplasmic tyrosine residue Y321 with phenylalanine (Y321F), the authors showed that the conserved Y321 was involved in the regulation of the expression of the AMPAR and the activation of key intracellular kinases, such as Fyn, Lyn, ERK, JNK and S6 kinase [
39]. At the same time, it was also shown that the Y321F substitution impacted synaptic structure, with Y321 playing a key role in maintaining excitatory synapses, which can affect synapse stability and function [
40]. These results are reminiscent of earlier investigations by our group showing that the replacement of Y320 (the equivalent in humans of mice Y321) by phenylalanine (Y320F) in a non-neuronal human cell line resulted in the abrogation of MHC-I endocytosis and the shedding of a pool of the open conformers into the extracellular milieu [
41], which could explain the above mentioned results as a consequence of impaired endocytosis and the subsequent intracellular trafficking. Importantly, both biochemical processes were associated with the phosphorylation of Y320 in the cytoplasmic tail of open MHC-I conformers [
42] (see ), most likely by the protein tyrosine kinase Lck. Indeed, these studies revealed that the phosphorylated (pY320) open HLA-I conformers showed an increased tendency to
cis-associate with cell surface receptors, such as CD8αβ and the transferrin receptor, and intracellular tyrosine kinases, such as Lck [
42].
Finally, a recent study by Saul Villeda’s group has shown that the H-2Kb allele, but not the H-2Db, plays a key role in the proliferation of neural stem and progenitor cells (NSPC) [
43]. Loss of H-2Kb, but not of H-2Db, increased NSPC proliferation. In contrast, overexpression of H-2Kb, but not of H-2Db, decreased proliferation. Noteworthy, additional experiments showed that H-2Kb regulates NSPC proliferation by inhibiting signaling through the fibroblast growth factor receptor (FGFR). These results are reminiscent of the studies of Michael Edidin and others showing the effect that MHC-I molecules have on the modulation of signal transduction events mediated by growth factor receptors, such as the IR and the EGFR, upon ligand binding [
8], and suggest that H-2Kb, but not H-2Db, may specifically
cis-associate with the FGFR and regulate the outside-in signaling.
5. Conclusions
Altogether, these results implicate both closed and open MHC-I conformers, as well as soluble MHC-I molecules and β2m, in the regulation of synaptic plasticity and neurogenesis and reinforce the view that the outside-in signals mediated by neuronal signaling receptors in humans (e.g., NMDAR, AMPAR, IR, FGFR, NCAM1, KIR, LILR,
etc.) that convey to the neuron are fine-tuned, at least in part, by MHC-I molecules. At this point, it is important to note that most of these conclusions have been obtained from studies where MHC-I molecules were absent or overexpressed, something rarely seen in homeostatic conditions. In contrast, very little data on the nature of the ligands involved in these results have been provided. Nevertheless, the accumulated evidence points to MHC class I molecules as modifiers of essential biochemical and biological processes that take place during brain homeostasis. Overall, it can be hypothesized that the non-immunological functions attributed to MHC-I molecules are likely linked to the fluctuation of the closed ↔︎ open conformers physiological equilibrium, which allows
cis-interactions with receptors for hormones and growth factors and trans-interactions with inhibitory receptors present in the cells of the CNS [
44,
45,
46], and to the existence of secreted forms of closed and open conformers in biological fluids, which can interact with their ligands expressed in neighboring cells. In either situation, MHC-I molecules will play a modulatory role in the signaling through those receptors.
Assuming that what happens in the immunological system as a result of
cis–
trans interactions between MHC-I molecules and activation/inhibitory receptors, namely at the level of NK and effector-memory CD8+ T cells, can be transposed to the cells of the CNS, it can be proposed that the
cis–
trans interactions of the closed and open MHC-I conformers with a diverse array of inhibitory and activating receptors expressed by neurons and glial cells, as well as receptors for hormones, growth factors, neurotransmitters, and others yet to be identified [
9,
13,
14,
44,
45,
46], will result in inhibition or activation of neuronal and other glial cells, resulting in modifications of neurogenesis, synaptic plasticity and density, synaptic pruning,
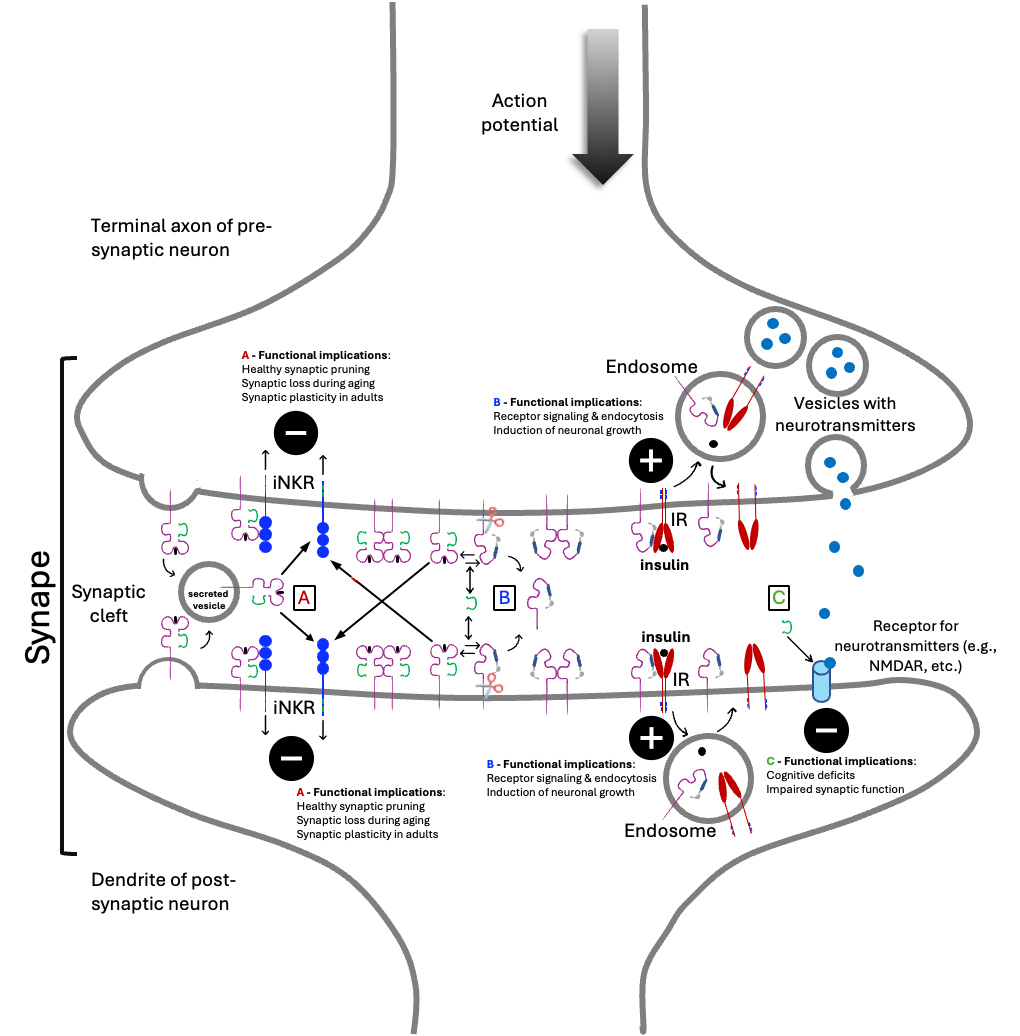
etc. The potential mechanisms of action of MHC-I molecules on neurons are schematized in , and the functional implications of these interactions are summarized in Box 1.
.
Summary of the Functional Implications of the cis–trans Interactions Between MHC-I Molecules and Their Receptors.
- Homeostatic expression of classical MHC-I molecules is critical for correct brain homeostasis and normal cognition and could function as a neuroprotective factor.
- MHC-I molecules regulate hippocampal synapse density by fine-tuning neuronal IR signaling, an effect that could be mediated by soluble MHC-I forms secreted by neurons.
- Endogenous MHC-I inhibits NMDAR function and controls AMPAR trafficking in the CNS.
- Soluble closed MHC-I conformers induce neuroinhibitory signals in vitro and are potential biomarkers of neurodegeneration in vivo.
- Cis-interactions between open MHC-I conformers and cell surface receptors favor neurogenesis.
- Trans-interactions between closed MHC-I conformers and cell surface receptors inhibit neuritogenesis.
- Absence of MHC-I molecules compromises the capacity of neurons to regenerate axons.
- Upregulation of MHC-I molecules after nerve transection positively influences neuroregeneration, which is influenced by the MHC-I haplotype both in mice and humans.
- An aberrant increase or decrease in MHC-I expression can disrupt synaptic plasticity and prompt cognitive deficits in the aging brain.
- An MHC-I expression threshold may settle the balance between neurodegeneration and neuroregeneration.
- The open/closed MHC-I conformers ratio seems critical for MHC-I signaling and function in neurons.
- Soluble β2m is an endogenous NMDAR antagonist that impairs synaptic function after aggregating with Aβ.
- β2m-Aβ aggregates are increased in the brains of AD patients and AD mice, and trigger neurodegeneration.
- Y321 (Y320 in humans) plays a key role in maintaining excitatory synapses, affecting synapse stability and function, and regulates the expression of the AMPAR and the activation of intracellular kinases.
- H-2Kb molecules regulate NSPC proliferation by inhibiting signaling through the FGFR.
. Scheme of potential mechanisms of action of MHC-I molecules on neurons based on scientific data from the Central Nervous System described in this article. There is evidence that the MHC-I regulated processes may occur in neurons from different areas (e.g., cortex, retina, hippocampus, spinal cord, <em>etc.</em>,) and both in excitatory (e.g., glutamatergic) or inhibitory (e.g., GABAergic) type of neurons. Glial cells, such as astrocytes and microglia, present in the surroundings of neuronal cells are also expected to be involved, but were not included for simplicity. The different forms of the classical MHC-I molecules are identical to the ones shown in . MHC-I molecules are reported to be expressed in both sides of the synapse. Monomeric closed conformers can dissociate generating monomeric open conformers, while releasing soluble β2m. Closed and open conformers can form homodimers, which ligands are unknown. Closed conformers can be secreted embedded in vesicles into the synaptic cleft as well as the open conformers, after cleavage by a metalloprotease (✂︎). Besides, open conformers can form heterodimers with receptors for hormones, growth factors, and others. In this model, all the events described can occur in the pre-synaptic (terminal axon) and post-synaptic (dendrite) neurons. The action potential of the pre-synaptic neurons and the vesicles containing neurotransmitters are indicated. Described <em>trans</em>-interactions of closed conformers with inhibitory NK receptors (iNKR) and the functional consequences are shown in <span style="color:#c00000;">A</span>. Described <em>cis</em>-interactions of open conformers with receptors for hormones and growth factors, such as the insulin receptor (IR), together with the functional implications are shown in <span style="color:#0432ff;">B. </span>The heterodimer is likely endocyted and the components recycled back. Described <em>trans-</em>interactions between β2m and open conformers or the glutamate receptor (NMDAR) and functional implications are indicated in <span style="color:#70ad47;">C</span>.
6. Future Perspectives
Brain homeostasis is complex and regulated by numerous factors besides MHC-I molecules. Nevertheless, based on the current knowledge, the fine-tuning of receptor signaling in brain cells by MHC-I molecules can be exerted by any of the possible players cited above and illustrated in . Based on the data presented, a series of strategies can be anticipated to modulate the balance between neurodegeneration and neuroregeneration towards the latter. These strategies must necessarily involve the modulation of the physiological equilibrium between the closed and open MHC-I conformers and identify the whole array of possible molecular partners. Most experimental studies point to the open conformers as positive and necessary for correct brain homeostasis, as an increase in the closed MHC-I conformers, caused by aging, inflammation, or high levels of IFN-γ, are all associated with neurodegeneration. Certainly, these strategies will only succeed if we can determine and quantitate the percentage of closed and open conformers that favors neuroregeneration and how to influence this balance. Regardless of the strategy, what is becoming certain is that closed and open conformers are truly gatekeepers of the good functioning of the brain and potential biomarkers of the onset of neurodegeneration.
Even though this opinion article focused mainly on neuronal cells, any future research will need to delve into the intricate interplay of circadian and cellular processes within the neuronal microenvironment. For instance, neuronal function is influenced by nearby glial cells, including astrocytes, microglia, oligodendrocytes,
etc., which play important roles in the control of pre- and post-synaptic activity and manage neurotransmitter dynamics [
47,
48]. More recent studies have pinpointed regions and cell types in the brain that are vulnerable to aging and defined hallmarks of the process, namely an increase in the immune responses and a decrease in neurogenesis, neuronal signaling and structural integrity [
49]. These data are in accord with studies of mice retinas, where glial cells seem to play an important role in modulating the immune response after insults to the retinal parenchyma by increasing MHC molecules [
50]. Even though these reports suggest that CD8+ T cell mediated responses could be harmful for neuronal integrity, the recent accumulated evidence, including our own, supports the opposite, that is, there may exist specific subpopulations of CD8+ T cells that are neuroprotective [
51,
52,
53,
54,
55,
56]. On the other hand, it has been reported that MHC-I expression is also strongly driven by oxidative stress [
57], highlighting the impact of age-related changes on nighttime pineal melatonin regulation, an antioxidant hormone that declines approximately tenfold between the second and ninth decades of life [
58]. This points to the possibility that age-related disruptions in how CNS and systemic cells reset and downregulate overnight could be key to understanding MHC-I regulation, including shifts between its open and closed conformations. Since astrocytes themselves produce melatonin [
59], any decline in this function with aging may impair the neuronal microenvironment’s ability to properly downregulate and reset [
60].
After all the above, it is possible to propose several therapeutic strategies that might provide novel insights into the role of MHC-I molecules and their ligands in the regulation of synaptic plasticity, neuronal survival and the balance between neurodegeneration and neuroregeneration (see Box 2). They only represent a few of the potential therapeutic strategies that might be developed.
.
Proposed Therapeutic Strategies to Regulate Synaptic Plasticity, Neuronal Survival and the Balance Between Neurodegeneration and Neuroregeneration.
- Downregulating increased levels of closed MHC-I conformers by using treatments that inhibit MHC-I expression on neuronal and glial cells might help reduce and/or prevent neuroinflammation, which often exacerbates neurodegenerative diseases like Alzheimer's, Parkinson's, and dementia. These may include cytokines, inhibitors of the proteasome or manipulation of epigenetic regulators of MHC-I expression.
- Increasing the levels of open MHC-I conformers by using treatments that induce the formation of open conformers, such as increasing metabolic activity, intracranial current stimulation, inducing the dissociation of β2m by cytokines, such as IL-15, or inducing phosphorylation of Y320. Any increase in open conformers on neuronal and glial cells might promote neuroregeneration.
- Blocking particular MHC-I-KIR/LILR interactions could shift the balance towards neuroregeneration. KIR and LILR are known to be involved in regulating microglial activity and neuronal cell death, and KIR/LILR overactivation can contribute to neurodegenerative processes. Indeed, certain interactions between MHC-I molecules and KIR/LILR in the brain could promote neuroinflammation.
- Enhancement of Insulin and Fibroblast Growth Factor Receptors signaling could promote neurogenesis and protect the brain from neurodegeneration. Indeed, the IR and FGFR are known to be involved in regulating neural cell growth, survival, and plasticity. Inhibition of IR signaling is also implicated in neurodegenerative diseases, such as Alzheimer's, where insulin resistance is a feature.
- Establishment personalized therapies might be necessary. Given the genetic variability in HLA-I allele expression and KIR/LILR receptor profiles among humans, studies of genetic or transcriptomic profiles of patients without and with neurodegenerative diseases could provide valuable information on possible biological targets.
Acknowledgments
This work is part of the M.Sc. thesis of Ricardo Carvalho.
Author Contributions
F.A.A. and R.A.S.C. wrote the manuscript; F.A.A. elaborated the figures; E.M.C. reviewed and edited the manuscript and figures.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
The writing of this opinion article received no external funding.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1.
Global Status Report on the Public Health Response to Dementia. Available online: https://www.who.int/publications/i/item/9789240033245 (accessed on 3 June 2023).
2.
Montealegre S, Van Endert PM. Endocytic Recycling of MHC Class I Molecules in Non-professional Antigen Presenting and Dendritic Cells.
Front. Immunol. 2019,
9, 3098. doi:10.3389/fimmu.2018.03098.
[Google Scholar]
3.
Vijayan S, Sidiq T, Yousuf S, Van Den Elsen PJ, Kobayashi KS. Class I transactivator, NLRC5: A central player in the MHC class I pathway and cancer immune surveillance.
Immunogenetics 2019,
71, 273–282. doi:10.1007/s00251-019-01106-z.
[Google Scholar]
4.
Jongsma MLM, Guarda G, Spaapen RM. The regulatory network behind MHC class I expression.
Mol. Immunol. 2019,
113, 16–21. doi:10.1016/j.molimm.2017.12.005.
[Google Scholar]
5.
Arosa FA, Santos SG, Powis SJ. Open conformers: the hidden face of MHC-I molecules.
Trends Immunol. 2007,
28, 115–123. doi:10.1016/j.it.2007.01.002.
[Google Scholar]
6.
Arosa FA, Esgalhado AJ, Reste-Ferreira D, Cardoso EM. Open MHC Class I Conformers: A Look through the Looking Glass.
Int. J. Mol. Sci. 2021,
22, 9738. doi:10.3390/ijms22189738.
[Google Scholar]
7.
Chiu SL, Chen CM, Cline HT. Insulin receptor signaling regulates synapse number, dendritic plasticity, and circuit function
in vivo.
Neuron 2008,
58, 708–719. doi:10.1016/j.neuron.2008.04.014.
[Google Scholar]
8.
Ramalingam TS, Chakrabarti A, Edidin M. Interaction of Class I Human Leukocyte Antigen (HLA-I) Molecules with Insulin Receptors and Its Effect on the Insulin-Signaling Cascade.
Mol. Biol. Cell. 1997,
8, 2463–2474. doi:10.1091/mbc.8.12.2463.
[Google Scholar]
9.
Dixon-Salazar TJ, Fourgeaud L, Tyler CM, Poole JR, Park JJ, Boulanger LM. MHC class I limits hippocampal synapse density by inhibiting neuronal insulin receptor signaling.
J. Neurosci. 2014,
34, 11844–11856. doi:10.1523/JNEUROSCI.4642-12.2014.
[Google Scholar]
10.
Held W, Mariuzza RA. Cis interactions of immunoreceptors with MHC and non-MHC ligands.
Nat. Rev. Immunol. 2008,
8, 269–278. doi:10.1038/nri2278.
[Google Scholar]
11.
Ruggiero FM, Springer S. Homotypic and heterotypic
in cis associations of MHC class I molecules at the cell surface.
Curr. Res. Immunol. 2022,
3, 85–99. doi:10.1016/j.crimmu.2022.05.001.
[Google Scholar]
12.
Adamashvili I, Kelley RE, Pressly T, McDonald JC. Soluble HLA: patterns of expression in normal subjects, autoimmune diseases, and transplant recipients.
Rheumatol. Int. 2005,
25, 491–500. doi:10.1007/s00296-005-0585-y.
[Google Scholar]
13.
Shatz CJ. MHC Class I: An Unexpected Role in Neuronal Plasticity.
Neuron 2009,
64, 40–45. doi:10.1016/j.neuron.2009.09.044.
[Google Scholar]
14.
Boulanger LM. Immune Proteins in Brain Development and Synaptic Plasticity.
Neuron 2009,
64, 93–109. doi:10.1016/j.neuron.2009.09.001.
[Google Scholar]
15.
Oliveira AL, Thams S, Lidman O, Piehl F, Hökfelt T, Kärre K, et al. A role for MHC class I molecules in synaptic plasticity and regeneration of neurons after axotomy.
Proc. Natl. Acad. Sci. USA 2004,
101, 17843–17848. doi:10.1073/pnas.0408154101.
[Google Scholar]
16.
Sabha M, Jr., Emirandetti A, Cullheim S, De Oliveira AL. MHC I expression and synaptic plasticity in different mice strains after axotomy.
Synapse 2008,
62, 137–148. doi:10.1002/syn.20475.
[Google Scholar]
17.
Capps GG, Robinson BE, Lewis KD, Zúñiga MC. In vivo dimeric association of class I MHC heavy chains. Possible relationship to class I MHC heavy chain-beta 2-microglobulin dissociation.
J. Immunol. 1993,
151, 159–169.
[Google Scholar]
18.
Smith JD, Myers NB, Gorka J, Hansen TH. Model for the in vivo assembly of nascent Ld class I molecules and for the expression of unfolded Ld molecules at the cell surface.
J. Exp. Med. 1993,
178, 2035–2046. doi:10.1084/jem.178.6.2035.
[Google Scholar]
19.
Jelonek MT, Classon BJ, Hudson PJ, Margulies DH. Direct binding of the MHC class I molecule H-2L
d to CD8: Interaction with the amino terminus of a mature cell surface protein.
J. Immunol. 1998, 160, 2809–2814.
[Google Scholar]
20.
Victório SC, Havton LA, Oliveira AL. Absence of IFNγ expression induces neuronal degeneration in the spinal cord of adult mice.
J Neuroinflammation. 2010,
7, 77. doi:10.1186/1742-2094-7-77.
[Google Scholar]
21.
Joseph MS, Bilousova T, Zdunowski S, Wu ZP, Middleton B, Boudzinskaia M, et al. Transgenic mice with enhanced neuronal major histocompatibility complex class I expression recover locomotor function better after spinal cord injury.
J. Neurosci. Res. 2011,
89, 365–372. doi:10.1002/jnr.22557.
[Google Scholar]
22.
Wootla B, Denic A, Watzlawik JO, Warrington AE, Zoecklein LJ, Papke-Norton LM, et al. Human class I major histocompatibility complex alleles determine central nervous system injury versus repair.
J. Neuroinflammation 2016,
13, 293. doi:10.1186/s12974-016-0759-4.
[Google Scholar]
23.
Tomiyama ALMR, Cartarozzi LP, de Oliveira Coser L, Chiarotto GB, Oliveira ALR. Neuroprotection by upregulation of the major histocompatibility complex class I (MHC I) in SOD1G93A mice. Front. Cell Neurosci. 2023, 17, 1211486. doi:10.3389/fncel.2023.1211486; Erratum in Front. Cell Neurosci. 2024,
18, 1493884. doi:10.3389/fncel.2024.1493884.
[Google Scholar]
24.
Zalocusky KA, Najm R, Taubes AL, Hao Y, Yoon SY, Koutsodendris N, et al. Neuronal ApoE upregulates MHC-I expression to drive selective neurodegeneration in Alzheimer’s disease.
Nat. Neurosci. 2021,
24, 786–798. doi:10.1038/s41593-021-00851-3.
[Google Scholar]
25.
Wu ZP, Washburn L, Bilousova TV, Boudzinskaia M, Escande-Beillard N, Querubin J, et al. Enhanced neuronal expression of major histocompatibility complex class I leads to aberrations in neurodevelopment and neurorepair.
J. Neuroimmunol. 2011,
232, 8–16. doi:10.1016/j.jneuroim.2010.09.009.
[Google Scholar]
26.
Needleman LA, Liu XB, El-Sabeawy F, Jones EG, McAllister AK. MHC class I molecules are present both pre- and postsynaptically in the visual cortex during postnatal development and in adulthood.
Proc. Natl. Acad. Sci. USA 2010,
107, 16999–17004. doi:10.1073/pnas.1006087107.
[Google Scholar]
27.
Glynn MW, Elmer BM, Garay PA, Liu XB, Needleman LA, El-Sabeawy F, et al. MHCI negatively regulates synapse density during the establishment of cortical connections.
Nat. Neurosci. 2011,
14, 442–451. doi:10.1038/nn.2764.
[Google Scholar]
28.
Kim MS, Cho K, Cho MH, Kim NY, Kim K, Kim DH, et al. Neuronal MHC-I complex is destabilized by amyloid-β and its implications in Alzheimer’s disease.
Cell Biosci. 2023,
13, 181. doi:10.1186/s13578-023-01132-1.
[Google Scholar]
29.
Irala D, Bonafina A, Fontanet PA, Alsina FC, Paratcha G, Ledda F. The GDNF-GFRα1 complex promotes the development of hippocampal dendritic arbors and spines via NCAM.
Development 2016,
143, 4224–4235. doi:10.1242/dev.140350.
[Google Scholar]
30.
Zhao Y, Zheng Q, Hong Y, Gao Y, Hu J, Lang M, et al. β2-Microglobulin coaggregates with Aβ and contributes to amyloid pathology and cognitive deficits in Alzheimer’s disease model mice.
Nat. Neurosci. 2023,
26, 1170–1184. doi:10.1038/s41593-023-01352-1.
[Google Scholar]
31.
Gao Y, Hong Y, Huang L, Zheng S, Zhang H, Wang S, et al. β2-microglobulin functions as an endogenous NMDAR antagonist to impair synaptic function.
Cell 2023,
186, 1026–1038.e20. doi:10.1016/j.cell.2023.01.021.
[Google Scholar]
32.
Escande-Beillard N, Washburn L, Zekzer D, Wu ZP, Eitan S, Ivkovic S, et al. Neurons preferentially respond to self-MHC class I allele products regardless of peptide presented.
J. Immunol. 2010,
184, 816–823. doi:10.4049/jimmunol.0902159.
[Google Scholar]
33.
Washburn LR, Zekzer D, Eitan S, Lu Y, Dang H, Middleton B, et al. A potential role for shed soluble major histocompatibility class I molecules as modulators of neurite outgrowth.
PLoS ONE 2011,
6, e18439. doi:10.1371/journal.pone.0018439.
[Google Scholar]
34.
Bilousova T, Dang H, Xu W, Gustafson S, Jin Y, Wickramasinghe L, et al. Major histocompatibility complex class I molecules modulate embryonic neuritogenesis and neuronal polarization.
J. Neuroimmunol. 2012,
247, 1–8. doi:10.1016/j.jneuroim.2012.03.008.
[Google Scholar]
35.
Cardoso EM, Lourenço-Gomes V, Esgalhado AJ, Reste-Ferreira D, Oliveira N, Amaral AS, et al. HLA-A23/HLA-A24 serotypes and dementia interaction in the elderly: Association with increased soluble HLA class I molecules in plasma.
HLA 2023,
102, 660–670. doi:10.1111/tan.15149.
[Google Scholar]
36.
Fourgeaud L, Davenport CM, Tyler CM, Cheng TT, Spencer MB, Boulanger LM. MHC class I modulates NMDA receptor function and AMPA receptor trafficking.
Proc. Natl. Acad. Sci. USA 2010,
107, 22278–22283. doi:10.1073/pnas.0914064107.
[Google Scholar]
37.
Nelson PA, Sage JR, Wood SC, Davenport CM, Anagnostaras SG, Boulanger LM. MHC class I immune proteins are critical for hippocampus-dependent memory and gate NMDAR-dependent hippocampal long-term depression.
Learn. Mem. 2013,
20, 505–517. doi:10.1101/lm.031351.113.
[Google Scholar]
38.
Frietze KK, Pappy AL 2nd, Melson JW, O’Driscoll EE, Tyler CM, Perlman DH, et al. Cryptic protein-protein interaction motifs in the cytoplasmic domain of MHCI proteins
. BMC Immunol. 2016,
17, 24. doi:10.1186/s12865-016-0154-z.
[Google Scholar]
39.
Eyford BA, Lazarczyk MJ, Choi KB, Varghese M, Arora H, Kari S, et al. Outside-in signaling through the major histocompatibility complex class-I cytoplasmic tail modulates glutamate receptor expression in neurons.
Sci. Rep. 2023,
13, 13079. doi:10.1038/s41598-023-38663-z.
[Google Scholar]
40.
Lazarczyk MJ, Eyford BA, Varghese M, Arora H, Munro L, Warda T, et al. The intracellular domain of major histocompatibility class-I proteins is essential for maintaining excitatory spine density and synaptic ultrastructure in the brain.
Sci. Rep. 2023,
13, 6448. doi:10.1038/s41598-023-30054-8.
[Google Scholar]
41.
Santos SG, Antoniou AN, Sampaio P, Powis SJ, Arosa FA. Lack of Tyrosine 320 Impairs Spontaneous Endocytosis and Enhances Release of HLA-B27 Molecules.
J. Immunol. 2006,
176, 2942–2949. doi:10.4049/jimmunol.176.5.2942.
[Google Scholar]
42.
Santos SG, Powis SJ, Arosa FA. Misfolding of Major Histocompatibility Complex Class I Molecules in Activated T Cells Allows cis-Interactions with Receptors and Signaling Molecules and Is Associated with Tyrosine Phosphorylation.
J. Biol. Chem. 2004,
279, 53062–53070. doi:10.1074/jbc.M408794200.
[Google Scholar]
43.
Lin K, Bieri G, Gontier G, Miller S, Smith LK, Snethlage CE, et al. MHC class I H2-Kb negatively regulates neural progenitor cell proliferation by inhibiting FGFR signaling.
PLoS Biol. 2021,
19, e3001311. doi:10.1371/journal.pbio.3001311.
[Google Scholar]
44.
Vidal GS, Djurisic M, Brown K, Sapp RW, Shatz CJ. Cell-autonomous regulation of dendritic spine density by PirB.
eNeuro 2016,
3, 1–15, doi:10.1523/ENEURO.0089-16.2016.
[Google Scholar]
45.
Djurisic M, Brott BK, Saw NL, Shamloo M, Shatz CJ. Activity-dependent modulation of hippocampal synaptic plasticity via PirB and endocannabinoids.
Mol. Psychiatry 2019,
24, 1206–1219. doi:10.1038/s41380-018-0034-4.
[Google Scholar]
46.
Djurišić M. Immune receptors and aging brain.
Biosci. Rep. 2024,
44, BSR20222267. doi:10.1042/BSR20222267.
[Google Scholar]
47.
Calì C, Cantando I, Veloz Castillo MF, Gonzalez L, Bezzi P. Metabolic Reprogramming of Astrocytes in Pathological Conditions: Implications for Neurodegenerative Diseases.
Int. J. Mol. Sci. 2024,
25, 8922. doi:10.3390/ijms25168922.
[Google Scholar]
48.
Kellogg CM, Pham K, Machalinski AH, Porter HL, Blankenship HE, Tooley KB, et al. Microglial MHC-I induction with aging and Alzheimer’s is conserved in mouse models and humans.
Geroscience 2023,
45, 3019–3043. doi:10.1007/s11357-023-00859-6.
[Google Scholar]
49.
Jin K, Yao Z, van Velthoven CTJ, Kaplan ES, Glattfelder K, Barlow ST, et al. Brain-wide cell-type-specific transcriptomic signatures of healthy ageing in mice.
Nature 2025,
638, 182–196. doi:10.1038/s41586-024-08350-8.
[Google Scholar]
50.
Intonti S, Kokona D, Zinkernagel MS, Enzmann V, Stein JV, Conedera FM. Glia Modulates Immune Responses in the Retina Through Distinct MHC Pathways.
Glia 2025,
73, 822–839. doi:10.1002/glia.24656.
[Google Scholar]
51.
Coder B, Wang W, Wang L, Wu Z, Zhuge Q, Su DM. Friend or foe: The dichotomous impact of T cells on neuro-de/re-generation during aging.
Oncotarget 2017,
8, 7116–7137. doi:10.18632/oncotarget.12572.
[Google Scholar]
52.
de Miranda, Zhang CJ, Katsumoto A, Teixeira AL. Hippocampal adult neurogenesis: Does the immune system matter?
J. Neurol. Sci. 2017,
372, 482–495. doi:10.1016/j.jns.2016.10.052.
[Google Scholar]
53.
Esgalhado AJ, Reste-Ferreira D, Albino SE, Sousa A, Amaral AP, Martinho A, et al. CD45RA, CD8β, and IFNγ Are Potential Immune Biomarkers of Human Cognitive Function.
Front. Immunol. 2020,
11, 592656. doi:10.3389/fimmu.2020.592656.
[Google Scholar]
54.
Zarif H, Nicolas S, Guyot M, Hosseiny S, Lazzari A, Canali MM, et al. CD8+ T cells are essential for the effects of enriched environment on hippocampus-dependent behaviour, hippocampal neurogenesis and synaptic plasticity.
Brain Behav. Immun. 2018,
69, 235–254. doi:10.1016/j.bbi.2017.11.016.
[Google Scholar]
55.
Levite M. Neuro faces of beneficial T cells: essential in brain, impaired in aging and neurological diseases, and activated functionally by neurotransmitters and neuropeptides.
Neural Regen. Res. 2023,
18, 1165–1178. doi:10.4103/1673-5374.357903.
[Google Scholar]
56.
Hu D, Weiner HL. Unraveling the dual nature of brain CD8+ T cells in Alzheimer’s disease.
Mol. Neurodegener. 2024,
19, 16. doi:10.1186/s13024-024-00706-y.
[Google Scholar]
57.
Anderson G, Almulla AF, Reiter RJ, Maes M. Redefining Autoimmune Disorders’ Pathoetiology: Implications for Mood and Psychotic Disorders’ Association with Neurodegenerative and Classical Autoimmune Disorders.
Cells 2023,
12, 1237. doi:10.3390/cells12091237.
[Google Scholar]
58.
Karasek M, Reiter RJ. Melatonin and aging.
Neuro Endocrinol. Lett. 2002,
23 (Suppl. 1), 14–16.
[Google Scholar]
59.
Liu YJ, Zhuang J, Zhu HY, Shen YX, Tan ZL, Zhou JN. Cultured rat cortical astrocytes synthesize melatonin: absence of a diurnal rhythm.
J. Pineal Res. 2007,
3, 232–238. doi:10.1111/j.1600-079X.2007.00466.x.
[Google Scholar]
60.
Anderson G. Physiological processes underpinning the ubiquitous benefits and interactions of melatonin, butyrate and green tea in neurodegenerative conditions.
Melatonin Res. 2024,
7, 20–46. doi:10.32794/mr112500167.
[Google Scholar]