1. Introduction
Freshwater fish are a vital component of freshwater ecosystems [
1], hold significant economic value [
2], and provide unique cultural value for residents [
3]. Unfortunately, due to climate change, dam construction, and other human activities, the habitat distribution, biological characteristics, and community composition of freshwater fish have been forced to change [
4,
5,
6,
7], with many species now on the brink of extinction. It is essential to conduct research on these species and implement conservation efforts to prevent their extinction. To address these challenges, integrating species distribution models with river network connectivity is a novel approach for habitat assessment of endangered species, which not only helps identify critical habitats but also provides insights into the impacts of human activities on freshwater ecosystems.
Schizothorax chongi (Fang, 1936;
S. chongi), belonging to the family Cyprinidae and the subfamily Schizothoracinae [
8], is an endemic fish species of the Upper Yangtze River Basin (UYRB). Due to overfishing, environmental degradation, and dam construction, the population of
S. chongi has been continuously declining [
9,
10]. Its wild population was officially listed as a National Class II protected species in China in 2021.
S. chongi is also an important economic fish species in its distribution area, known for its tender flesh, delicious flavor, and high market price [
9]. Therefore, conducting research and implementing conservation measures for
S. chongi is of paramount significance. Currently, research on
S. chongi primarily focuses on developmental and growth aspects [
7,
11,
12], the influence of environmental factors on swimming and spawning [
11,
12,
13,
14,
15], aquaculture techniques and disease management [
16,
17,
18], population dynamics and ecology [
19,
20], genetics and nutrition [
9,
21,
22], as well as anesthesia effect experimentation on juvenile fish [
23]. However, there is a lack of assessments regarding its potential habitat distribution and connectivity.
Species Distribution Models (SDMs) are useful tools for identifying potential habitat distributions for species [
24] and are increasingly applied in freshwater ecosystems [
25,
26,
27] as well as for freshwater fish [
28]. Biologists and managers can utilize SDMs to identify critical habitats, prioritize the locations of protected areas, and appropriately relocate endangered species [
24]. Therefore, it is feasible and necessary to employ SDMs to assess the potential habitat distribution of
S. chongi. River network connectivity (RNC) is defined as the water-mediated transfer of matter, energy, and organisms along longitudinal (upstream-downstream), lateral, and vertical dimensions [
29,
30]. For centuries, dam construction has disrupted RNC [
31,
32], significantly altering hydrological conditions and material flow processes in rivers [
29,
33], thus threatening the dispersal and migration of freshwater fish, which are critical life history processes [
34,
35]. Amid the global surge in dam construction, quantifying and assessing RNC and its changes is crucial for the conservation of fluvial fish diversity. The Dendritic Connection Index (DCI) [
36] is an effective tool for quantifying the impacts of dam construction on RNC, as it implicitly considers the movement and arrival probabilities of organisms within the river network. Therefore, when implementing conservation measures, in addition to considering the distribution of habitat suitability, river connectivity needs to be considered.
To address the knowledge gap of
S. chongi, we employed SDMs and the DCI [
36] to assess the potential habitat distribution and river network connectivity of
S. chongi during the periods 1970–2000 and 2001–2020 (2000: dividing line for whether dams in the UYRB were completed on a large scale) [
37]. This study can provide concrete recommendations for the restoration of
S. chongi habitats in the current UYRB and offer valuable references for researchers in freshwater fish conservation.
2. Materials and Methods
2.1. Study Area
Upper Yangtze River Basin (UYRB), located between 97.62°–110.30° E and 21.22°–34.55° N, covers an area of approximately 1 ×10
6 km
2 and extends a total length of 4511 km. This basin features significant topographical variation, with an elevation difference reaching 6790 m. The annual average temperature in the UYRB ranges from 8.6 to 16.8 °C, with the average temperature in the coldest month ranging from −16 to 15 °C. Annual precipitation ranges from 723 to 1134 mm, displaying a spatial distribution characterized by lower amounts in the west and higher amounts in the east, with uneven seasonal distribution mainly concentrated between April and October. Influenced by the East Asian monsoon, South Asian monsoon, and the topography of the Tibetan Plateau, the UYRB exhibits distinct regional climatic characteristics, primarily impacted by climate disasters such as low temperatures, droughts, and floods.
2.2. Species Occurrence
To collect as much data as we can, occurrence data for
S. chongi was primarily obtained from two sources: (1) through literature surveys (CNKI search keyword:
Schizothorax chongi; in Chinese) (https://www.cnki.net/; assessed on 9 June 2023) and (2) sampling conducted by the Institute of Hydrobiology, Chinese Academy of Sciences (listed in Supporting Information). All occurrence records were checked for synonyms and naming errors, and their spatial accuracy was verified before modeling [
38], ensuring that all occurrence points fell within the river network of the UYRB. To eliminate the effects of spatial autocorrelation, the occurrence records were thinned [
39,
40], retaining only one record per grid cell at a spatial resolution of 30 arc seconds [
41]. After processing, a total of 26 and 24 occurrence records for
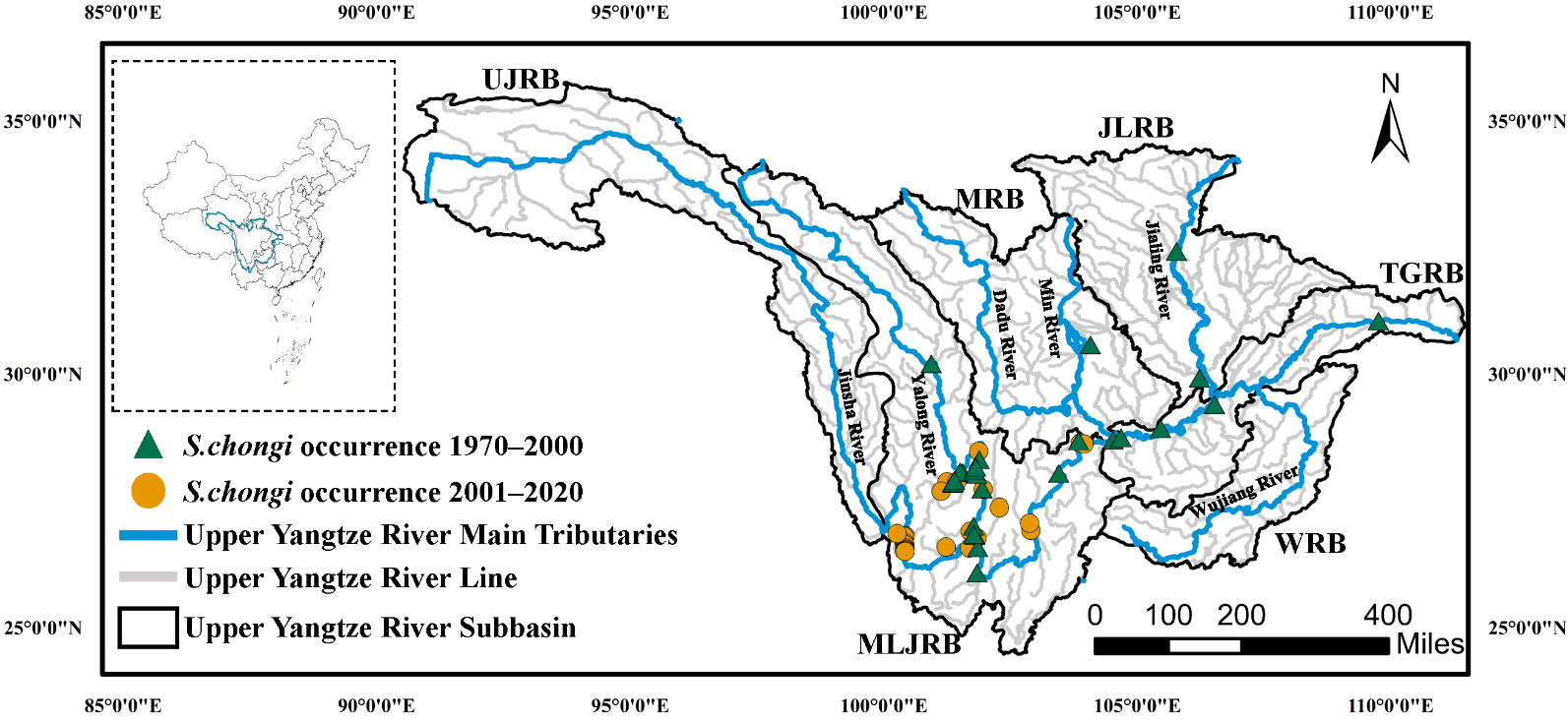
S. chongi were retained for modeling for the periods 1970–2000 and 2001–2020, respectively ().
. The location of the study area and the occurrence of <em>S. chongi </em>in the periods of 1970–2000 and 2001–2020. The Upper Yangtze River Basin is divided into five major sub-basins (): the Upper Jinsha River Basin (UJRB), the Middle and Lower Jinsha River Basin (MLJRB), the Min River Basin (MRB), the Jialing River Basin (JLRB), and the Wu River Basin (WRB), as well as the Three Gorges Reservoir Basin (TGRB).
We considered the impact of environmental factors as Zhang et al. (2020) [
42] suggested and selected a total of 25 environmental variables as candidate modeling factors including 19 climate variables and six human activity-related variables ().
.
Pre-selected modelling factor descriptions and sources (All the datas here were assessed on 13 March 2023).
| Type |
Variables |
Description |
Source |
|
BIO1 |
Annual Mean Temperature |
https://www.worldclim.org/data/worldclim21.html |
|
BIO2 |
Mean Diurnal Range |
https://www.worldclim.org/data/worldclim21.html |
|
BIO3 |
Isothermality |
https://www.worldclim.org/data/worldclim21.html |
|
BIO4 |
Temperature Seasonality |
https://www.worldclim.org/data/worldclim21.html |
|
BIO5 |
Max Temperature of Warmest Month |
https://www.worldclim.org/data/worldclim21.html |
|
BIO6 |
Min Temperature of Coldest Month |
https://www.worldclim.org/data/worldclim21.html |
|
BIO7 |
Temperature Annual Range |
https://www.worldclim.org/data/worldclim21.html |
|
BIO8 |
Mean Temperature of Wettest Quarter |
https://www.worldclim.org/data/worldclim21.html |
|
BIO9 |
Mean Temperature of Driest Quarter |
https://www.worldclim.org/data/worldclim21.html |
| Climate |
BIO10 |
Mean Temperature of Warmest Quarter |
https://www.worldclim.org/data/worldclim21.html |
|
BIO11 |
Mean Temperature of Coldest Quarter |
https://www.worldclim.org/data/worldclim21.html |
|
BIO12 |
Annual Precipitation |
https://www.worldclim.org/data/worldclim21.html |
|
BIO13 |
Precipitation of Wettest Month |
https://www.worldclim.org/data/worldclim21.html |
|
BIO14 |
Precipitation of Driest Month |
https://www.worldclim.org/data/worldclim21.html |
|
BIO15 |
Precipitation Seasonality |
https://www.worldclim.org/data/worldclim21.html |
|
BIO16 |
Precipitation of Wettest Quarter |
https://www.worldclim.org/data/worldclim21.html |
|
BIO17 |
Precipitation of Driest Quarter |
https://www.worldclim.org/data/worldclim21.html |
|
BIO18 |
Precipitation of Warmest Quarter |
https://www.worldclim.org/data/worldclim21.html |
|
BIO19 |
Precipitation of Coldest Quarter |
https://www.worldclim.org/data/worldclim21.html |
|
CLA |
Crop Land Area |
https://sedac.ciesin.columbia.edu/data/set/aglands-croplands-2000 |
|
DPI |
Development Pressure Index |
https://sedac.ciesin.columbia.edu/data/set/lulc-development-threat-index |
| Human |
HIP |
The Human Impervious Area Percentage |
https://sedac.ciesin.columbia.edu/data/set/ulandsat-hbase-v1 |
| Activity |
POPC |
Population Count |
https://sedac.ciesin.columbia.edu/data/set/gpw-v4-population-count-rev11 |
|
POPD |
Population Density |
http://sedac.ciesin.columbia.edu/data/set/gpw-v3-population-density |
|
HII |
The Human Influence Index |
http://sedac.ciesin.columbia.edu/data/set/wildareas-v2-human-influence-index-geographic |
The climate variables were derived from the WorldClim database (http://www.worldclim.org/; assessed on 13 March 2023), which provides commonly used bioclimatic variables for ecological niche modeling (BIO1–BIO19) [
43]. These data were downscaled from CRU-TS-4.06 [
44] by the Climate Research Unit at the University of East Anglia and bias-corrected using WorldClim 2.1 [
45]. The resolution of the climate variables is 30 arc seconds, and all six human activity-related factors (specific references can be found in Table A1) were resampled to the same resolution before selection.
We selected the modeling variables in two steps. First, we conducted modeling using the candidate factors, sequentially removing the climate variables related to temperature and precipitation, andthe human-related variables with low contributions, ultimately retaining two climate variables and two human activity-related variables. Next, we performed a collinearity test on the four selected variables by calculating their Variance Inflation Factor (VIF) values to ensure that all VIF values were less than 10, thus avoiding overfitting and eliminating the effects of multicollinearity [
46].
2.4. Ensemble Modeling
Ensemble modeling, by integrating the strengths of multiple models, mitigates the limitations inherent in single-model approaches, thereby enhancing the accuracy, stability, and reliability of predictions [
47]. In this study, we selected four algorithms to establish the base submodels required for the ensemble model (weighted by AUC; see below): Artificial Neural Networks (ANN) [
48], Surface Range Envelope Models (SRE) [
49], Flexible Discriminant Analysis (FDA) [
50], and Maximum Entropy Models (MAXENT) [
51], as these algorithms perform well and are not prone to overfitting. All algorithms were fitted using the default settings in the biomod2 package [
52] within the open-source statistical software R 4.3.1. The models were evaluated on the assessment dataset (20~30% of the combined occurrence and pseudo-absence data) using the Area Under the Receiver Operating Characteristic (ROC) Curve (AUC) [
53] and the True Skill Statistic (TSS) [
54]. The ROC curve illustrates the relationship between the true positive rate and the false positive rate at different thresholds. A ROC curve that bends more toward the upper left corner indicates a higher AUC value, which represents better model predictive performance. The True Skill Statistic (TSS) quantifies model performance, calculated as TSS = specificity + sensitivity − 1. A higher TSS value signifies better model predictive performance. Generally, when the AUC is greater than 0.90 and the TSS is greater than 0.75, the model performance is considered excellent [
55,
56]. Subsequently, this study employed repeated split-sampling and cross-validation to evaluate the models, retaining only those with an AUC of 0.75 to create an ensemble model weighted by AUC [
57,
58]. We randomly selected 70% of the total occurrence and pseudo-absence (randomly generated in grids without occurrence, with the same number as the occurrence) records as the training set for fitting the algorithms, while the remaining 30% were reserved for assessing algorithm performance. This process was repeated 10 times to minimize biases in dataset splitting caused by the variability of individual algorithms and to enhance the robustness of the results.
2.5. DCI Calculation
The physical RNC was evaluated using the DCI, which identifies river connectivity as a layered system operating across the entire network and considers stream segments as essential parts of a river continuum extending from its source to its mouth [
36]. It assesses the extent to which river connectivity is disrupted by barriers such as dams, waterfalls, locks, and weirs by estimating the likelihood of a fish successfully crossing these obstacles to travel between two segments within a river network [
36]. The DCI is determined using factors such as the number of barriers, their spatial distribution, biological passability, and river length, with scores ranging from 0 (wholly disconnected) to 100 (fully connected). It can be applied to calculate connectivity for potamodromous fish migrating within freshwater systems and diadromous fish moving between freshwater and ocean environments. On the assumption that the pass rates between dams are mutually independent, the formula for calculating the DCI for potamodromous fish is as follows:
where
n is the total number of the sections of the river network;
i and
j are the river section numbers;
L is the total length of the river network; $$l_{i} $$ and $$l_{j}$$ are the lengths of sections
i and
j, respectively; $$C_{i j}$$ is passability that refers to the probability of fish being able to cross a barrier in both the upstream or downstream direction, which is calculated as
where
M is the number of the dams between sections
i and
j; $$P_m^U$$ and $$P_m^D$$ are the upstream and downstream passibility of the
mth dam, respectively, which is assumed to have equal possibility to move upstream and downstream for potamodromous fish [
59].
To calculate the DCI for the UYRB, we began by verifying the geographical location of the dams to confirm their alignment with the river network. Next, all the dams within the basin were classified into three categories based on their construction or planned construction dates: pre-2000, post-2000, and those constructed between these two periods. Finally, the river networks were divided into segments according to the geographical locations of the dams in the three time periods, which were then used to calculate the DCI. The DCI was calculated using the Fish Passage Extension (FIPEX) for ArcGIS 10.4 on the ArcGIS platform [
60]. To calculate the DCI, the probability
P in Equation (2) must be determined. Passability is influenced by various factors, including physical characteristics (e.g., dam height, flow rate, and hydrodynamics), chemical conditions (e.g., pollution), and the biology of the species involved, which can vary by species, age, and other factors [
36]. In this study, only the impact of dam height was considered, with a fixed pass rate assigned to dams of different heights (). For dams taller than 70 m, classified as large dams by the International Commission on Large Dams, the pass rate is set to 0. For dams exceeding 15 m in height, the passability value is set to 0.6.
.
Dam height and corresponding pass rate.
| Dam Height (m) |
Pass Rate |
| <15 |
0.6 |
| 15–30 |
0.4 |
| 30–70 |
0.2 |
| >70 |
0 |
3. Results
3.1. Model Performance and Predominant Predictors
The habitat models for
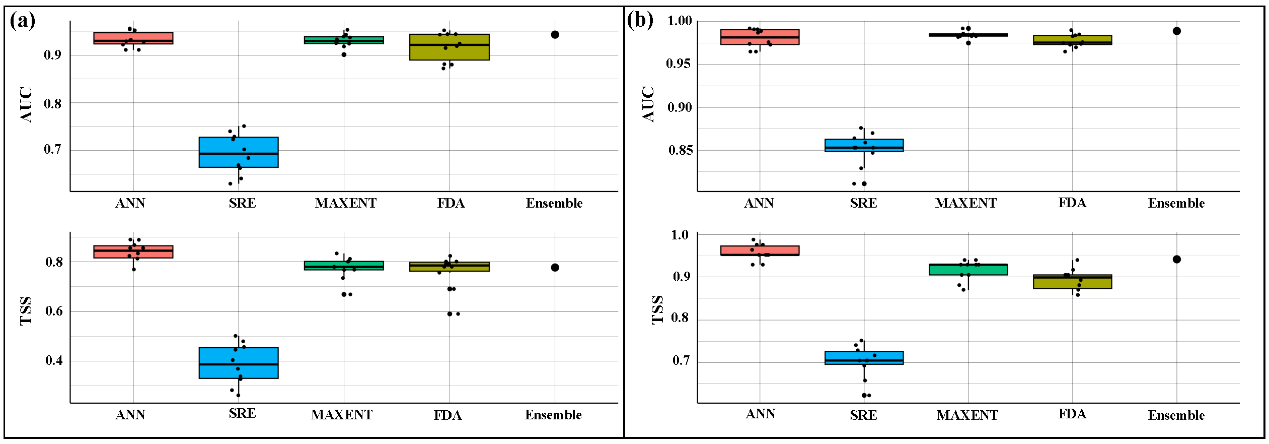
S. chongi during the two periods (1970–2000 and 2001–2020) were successfully established, demonstrating excellent model performance. The ensemble model for the 1970–2000 period achieved an AUC of 0.943 and a TSS of 0.777, while the habitat model for the 2001–2020 period achieved an AUC of 0.989 and a TSS of 0.942 (). Both AUC and TSS values exceed 0.85 and 0.75, respectively, indicating that our models are accurate and reliable.
After the selection process, the four retained modeling factors in this study were BIO7 (temperature annual range), BIO11 (mean temperature of coldest quarter), CLA (crop land area), and POPD (population density). The modeling factors for both the 1970–2000 and 2001–2020 periods underwent collinearity testing, and their model contributions and VIF values are presented in .
.
Relative contribution (RC) and Variance inflation factor (VIF) of modelling factor in 1970–2000 and 2001–2020.
| Modelling Periods |
Variables |
RC |
VIF |
| 1970–2000 |
BIO7 |
18.7% |
6.405106 |
| BIO11 |
54.0% |
7.871423 |
| CLA |
12.9% |
2.479721 |
| POPD |
14.4% |
2.275276 |
| 2001–2020 |
BIO7 |
35.2% |
6.165412 |
| BIO11 |
33.8% |
7.818387 |
| CLA |
12.9% |
2.364912 |
| POPD |
18.1% |
1.953089 |
. Model performance of sub-models and ensemble model (AUC and TSS) for <em>S. chongi</em> in (<strong>a</strong>) 1970–2000 and (<strong>b</strong>) 2001–2020; AUC means Area Under the Curve and TSS means True Skill Statistic.
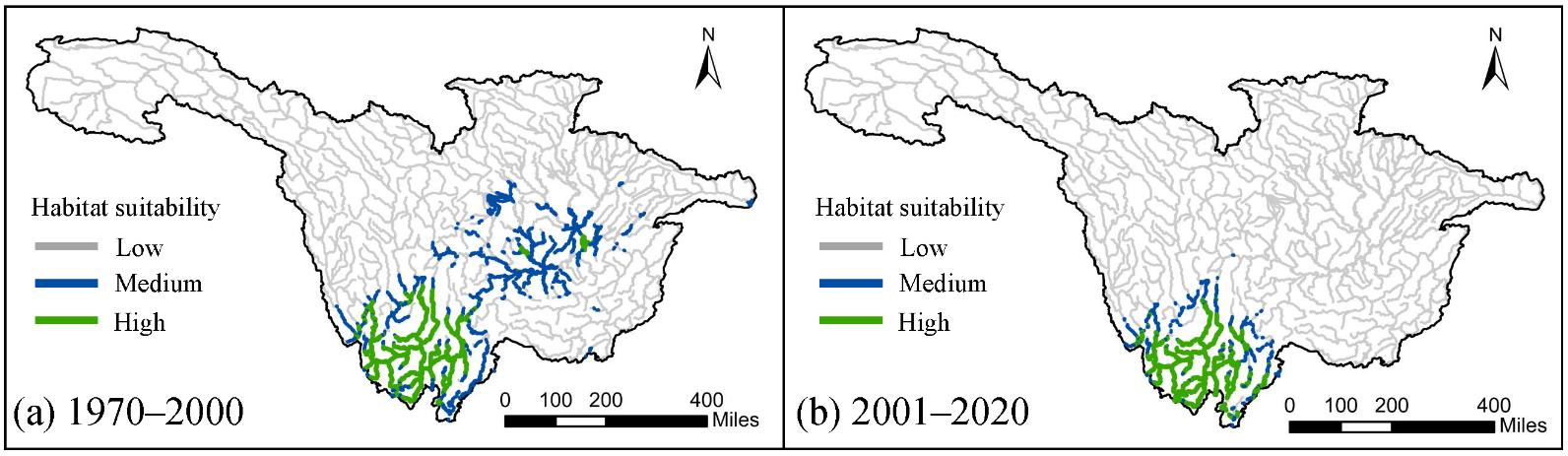
The simulated potential habitat distribution of
S. chongi for the periods 1970–2000 and 2001–2020 is shown in .
S. chongi experienced a significant reduction in habitat, with the lengths of medium and high suitability habitats decreasing from 10,213.76 km in the 1970–2000 period to 4932.04 km in the 2001–2020 period, representing a decline of 51.7%. The predominant type of habitat reduction was medium suitability, which decreased by 4711.78 km, primarily occurring outside the Jinsha River basin. Notably, there was no significant reduction in high suitability habitats in the middle and lower reaches of the Jinsha River between the two periods ().
. Habitat suitability distribution for <em>S. chongi</em> in 1970–2000 and 2001–2020. It was categorized into three levels: Low (habitat suitability < 0.4), Medium (habitat suitability 0.4–0.7), and High (habitat suitability 0.7–1).
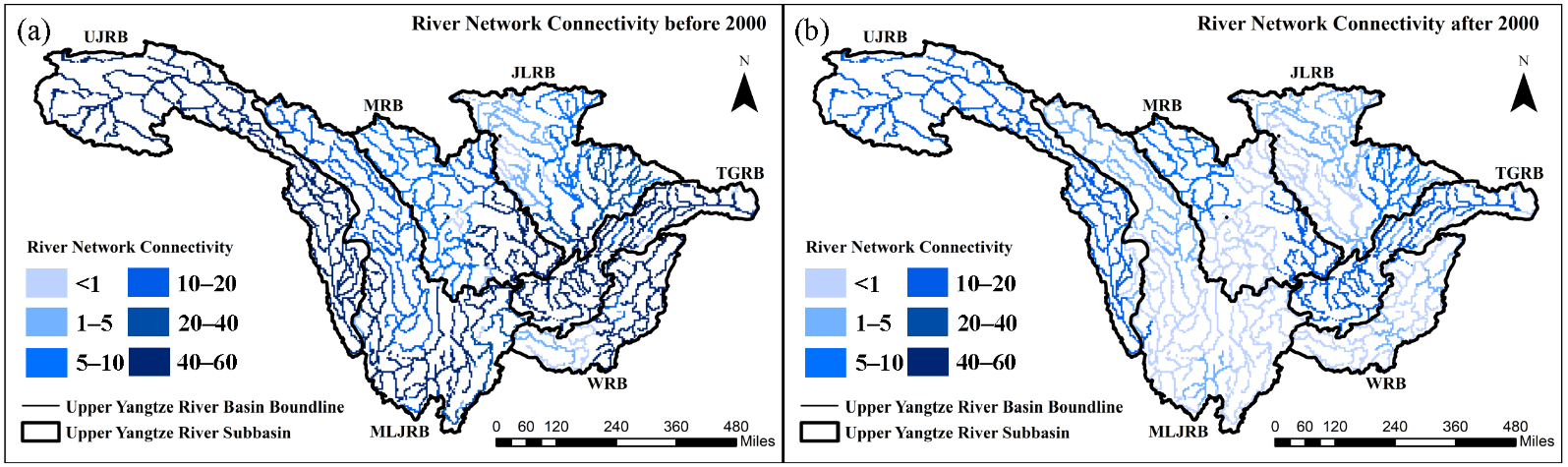
Comparisons of DCI distribution in different periods show that the RNC experienced a drastic reduction with the continuous advancement of hydropower development in the UYRB (). Before 2000, although some dams had been constructed in the UYRB, the DCI values of the mainstream still ranged between 40 and 60. Most of the sub-basins had DCI values above 10, with the exception of the northern part of the Jialing River, where the DCI was below 10. After 2000, the national government began to heavily develop the hydropower resources of the UYRB, resulting in a significant decline in the DCI values across the entire basin, with some parts of the mainstream in the MLJRB having DCI values below 1. Additionally, the DCI in the MRB and WRB also saw substantial declines, with more than 50% of these areas having DCI values below 1. Only the headwaters of UJRB, TGRB, and their surrounding areas maintained relatively high DCI values (greater than 10).
. The distribution of river network connectivity of the Upper Yangtze River Basin (<strong>a</strong>) before and (<strong>b</strong>) after 2000.
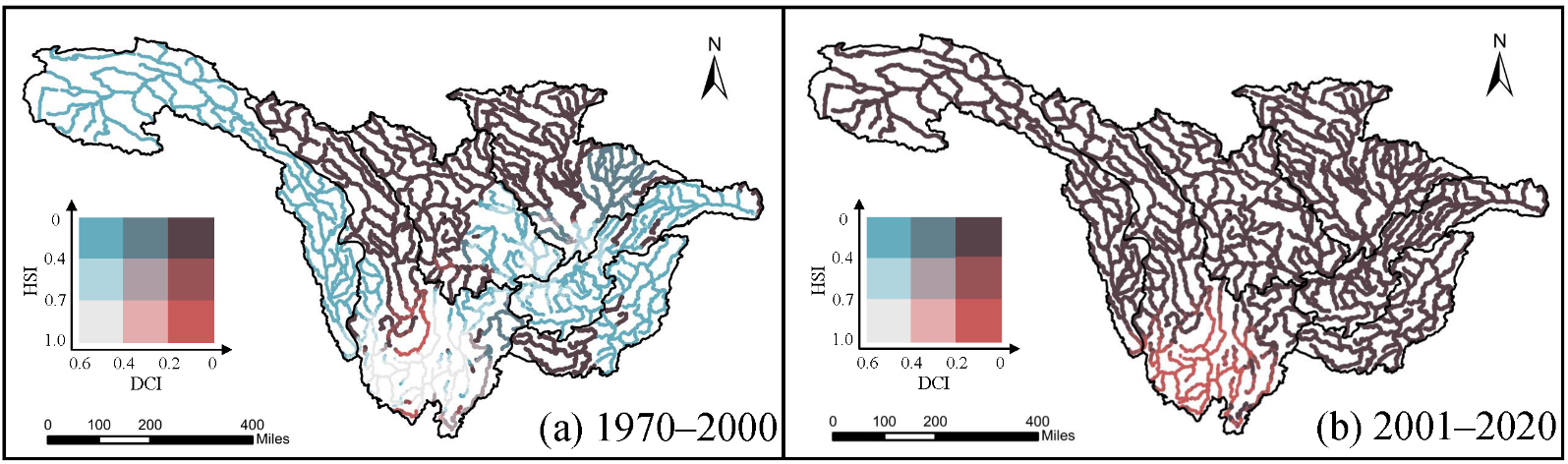
The bivariate map of the Habitat Suitability Index (HSI) and the Dendritic Connectivity Index (DCI) for
S. chongi during the periods 1970–2000 and 2001–2020 are illustrated in . In the 1970–2000 period, the southern region of the Middle and Lower Jinsha River Basin (MLJRB) exhibited both high connectivity and high suitability, while a region at the junction of the Min River, Jialing River, and Three Gorges Basin showed high connectivity and medium suitability. However, in the 2001–2020 period, connectivity and habitat suitability in the junction area of the Min River, Jialing River, and Three Gorges Basin declined simultaneously, resulting in a complete loss of colonization potential in this area. Furthermore, although the southern region of the MLJRB maintained relatively high habitat suitability, its connectivity decreased.
. Bivariate distribution of Dendritic Connectivity Index (DCI) and Habitat Suitability Index (HSI) in periods (<strong>a</strong>) 1970–2000 and (<strong>b</strong>) 2001–2020 for <em>S. chongi</em>.
4. Discussion
Our study assessed the suitability and connectivity of
S. chongi habitat for 1970–2000 and 2001–2020 from two critical perspectives. The results reveal a significant decline in both river network connectivity in the Upper Yangtze River Basin (UYRB) and habitat suitability for
S. chongi, serving as a further warning for the conservation of the wild population of this valuable economic fish species. This research is the first to provide a basin-scale assessment of the potential habitat distribution and connectivity of
S. chongi, which can assist local governments in formulating specific conservation measures and plans.
Overall, the modeling results for 1970–2000 and 2001–2020 indicate that BIO7 (temperature annual range) and BIO11 (mean temperature of coldest quarter) are the most significant contributing factors in the models (). BIO7 and BIO11 are temperature-related climate factors, and air temperature is often highly correlated with water temperature [
61]. As a cold-water species [
13],
S. chongi is reasonably sensitive to changes in water temperature. CLA (crop land area) and POPD (population density) are the human activity-related factors selected. CLA has been shown to negatively impact the survival of freshwater fish [
62], and high levels of POPD can exacerbate the pressures faced by freshwater fish populations [
63], such as pollution of the aquatic environment, night time light pollution, the construction of more barriers, and fishing activities [
64].
During the period from 2001 to 2020, only the habitat of
S. chongi in the southern region of the Middle and Lower Jinsha River Basin (MLJRB) was preserved (), which is consistent with the distribution of occurrence points (). Furthermore, the connectivity of the entire Upper Yangtze River Basin (UYRB) has declined, posing a threat to the remaining habitat of
S. chongi in MLJRB (). Fortunately, since dam construction plans often start from the downstream of tributaries, the headwaters of UJRB, TGRB, and their surrounding areas maintained relatively high connectivity with only a slight decrease. These results provide strong evidence supporting Han et al. (2009) and Duan et al. (2010) [
9,
10], indicating a deterioration in the survival conditions of
S. chongi and a continuous decline in its resource availability.
Conserving biodiversity requires the formulation of appropriate strategies [
65], particularly in the Upper Yangtze River Basin (UYRB), where the diversity of freshwater fish and the number of related conservation areas are extremely unbalanced [
66]. This study simultaneously assesses river network connectivity and habitat suitability (ecological factors), which aids in effective river conservation [
67,
68]. For
S. chongi, the most urgent task is to restore river connectivity in the southern Middle and Lower Jinsha River Basin (MLJRB), as habitat suitability here is still high, to protect the last remaining habitats of its wild population. For instance, fish passages specifically designed for
S. chongi can be constructed to facilitate their upstream migration in the MLJRB. Additionally, other measures can be implemented. On one hand, artificial breeding and release programs upstream of dams can be employed to enhance the population size of wild individuals. On the other hand, optimizing dam operations with greater consideration for ecological benefits is also essential.
In the calculation of Dendritic Connectivity Index (DCI), due to the lack of measured data for specific fish species, we assumed a fixed pass rate for the fish [
69], which might lead to an incorrect assessment of the fish's migratory ability. Additionally, this study assumes that the ability of fish to pass a particular dam is independent of their ability to pass other dams, which may not hold true in many cases [
69]. Further research is needed to address the issue of biological passability from both physical (flow regulation) and biological (species physiology) perspectives [
36]. Therefore, the DCI calculated in this study may deviate from the actual situation of the fish. At smaller spatial scales, the distribution of species may be shaped by both abiotic factors (such as flow dynamics and hydrological conditions) and biotic factors (including interspecies relationships and resource availability) [
70]. While our models focus on broad-scale patterns and project potential habitats, these simulations may not fully align with local-scale habitat suitability [
37]. Further research could enhance accuracy by incorporating fine-scale environmental variables, such as river morphology, water chemistry, and ecological interactions, to better reflect actual habitat distributions.
5. Conclusions
This study utilized the Dendritic Connectivity Index (DCI) and Species Distribution Models (SDMs) to assess the habitat connectivity and suitability of S. chongi during the periods of 1970–2000 and 2001–2020. The results indicate that from 2001 to 2020, there was a substantial reduction in the habitat of S. chongi, with a total length of medium and high suitability habitats decreasing by 51.7%. The high suitability habitat in the southern section of the Middle and Lower Jinsha River Basin (MLJRB) did not experience a significant reduction. Although this region maintained relatively high habitat suitability during 2001–2020, the connectivity has significantly declined, making the restoration of connectivity in this area extremely urgent. Implementing fish stocking upstream of the dam and constructing fish passages in the MLJRB are effective restoration measures. The findings provide effective recommendations for conserving both the current and future wild populations of S. chongi.
Appendix A
Table A1.
Sources of human activity related variables.
| Variables |
Description |
| CLA |
Crop Land Area [71,72] |
| DPI |
Development Pressure Index [73,74] |
| HIP |
The Human Impervious Area Percentage [75] |
| POPC |
Population Count [76] |
| POPD |
Population Density [77] |
| HII |
The Human Influence Index [78] |
Acknowledgments
We especially thank Zhi Yang for providing fish occurrence data for this study.
Author Contributions
Conceptualization, P.Z.; Methodology, P.Z.; Software, X.B.; Data Curation, X.B., Y.Y., M.X. and L.Q.; Writing–Original Draft Preparation, X.B.; Writing–Review & Editing, X.B. and P.Z.; Visualization, X.B. and Z.N.; Supervision, P.Z.
Ethics Statement
This study does not involve human or animal subjects.
Informed Consent Statement
Not applicable.
Data Availability Statement
Original data and code are available from the corresponding author upon reasonable request.
Funding
This study was financially supported by the National Natural Science Foundation of China (Nos.52179142) and the Key R&D Program of Hubei Province (No. 2023BCB110).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1.
Lundberg JG, Kottelat M, Smith GR, Stiassny ML, Gill AC. So many fishes, so little time: an overview of recent ichthyological discovery in continental waters.
Ann. Mo. Bot. Gard. 2020,
87, 26–62.
[Google Scholar]
2.
Nelson JS, Grande TC, Wilson MV. Fishes of the World, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016. ISBN: 9871118342336.
3.
Reid GM, Contreras MacBeath T, Csatádi K. Freshwater Fishes and Their Conservation: Global Challenges.
Int. Zoo Yb. 2013,
47, 6–45.
[Google Scholar]
4.
Parmesan C, Yohe G. A globally coherent fingerprint of climate change impacts across natural systems.
Nature 2003,
421, 37–42.
[Google Scholar]
5.
Ziv G, Baran E, Nam S, Rodríguez-Iturbe I, Levin SA. Trading-off fish biodiversity, food security, and hydropower in the Mekong River Basin.
Proc. Natl. Acad. Sci. USA 2012, 109, 5609–5614.
[Google Scholar]
6.
Dudgeon D, Arthington AH, Gessner MO, Kawabata Z, Knowler DJ, Lévêque C, et al. Freshwater biodiversity: Importance, threats, status and conservation challenges.
Biol. Rev. Camb. Philos. Soc. 2006,
81, 163–182.
[Google Scholar]
7.
Cheng XY, Tao J, Wu RD, Chen LQ, Ding CZ. Functional ecology of freshwater fish: research progress and prospects.
Acta Ecol. Sin. 2019,
39, 810–822. (In Chinese)
[Google Scholar]
8.
Chen LQ, Wu Q, Zheng SM, Liu FJ. Development of embryo and yolk-sac larva of
Schizothorax chongi.
J. Fish. Sci. China 2008,
15, 927–934. (In Chinese)
[Google Scholar]
9.
Duan B, Liu HY. Study on Isozymic Tissue-Specificity in
Schizothorax chongi Fang.
J. Southwest Univ. (Nat. Sci.Ed.) 2010,
32, 27–30. (In Chinese)
[Google Scholar]
10.
Han JC, Liu GY, Zhuge YS, Huang YP, Liu DF. Ecological Protection of Fishery Resources Based on Development of Hydropower in Yalona River.
J. China Three Gorges Univ. (Nat. Sci.Ed.) 2009,
31, 15–19. (In Chinese)
[Google Scholar]
11.
Qin XH, Liu GY, Wu YJ, Shi XT Wang XL. Effects of Light Intensity on the Hatching Rate of Fertilized
Schizothorax chongi Eggs and on the Growth and Feeding of Larvae.
J. Hydroecol. 2017,
38, 97–102. (In Chinese)
[Google Scholar]
12.
Zhu QG, Tang HY, Lin H, Gong Y, Li XN, Yang Z. Age Structure, Growth Characteristics and Population Dynamic of
Schizothorax chongi in Middle and Lower Jinsha River.
J. Hydroecol. 2021,
42, 56–63. (In Chinese)
[Google Scholar]
13.
Liu GY, Han JC, Tu ZY, Ma ZK, Shi XT, Huang YP. Influence of Water Velocity on Hematology Indices of
Schizothorax chongi.
J. Anhui Agric. Sci. 2011,
39, 7298–7300. (In Chinese)
[Google Scholar]
14.
Yuan X, Tu ZY, Han JC, Wang XX, Shi XT, Liu GY, et al. Effects of Flow Rate on Swimming Behavior and Energy Consumption of Schizothorax chongi.
Acta Hydrobiol. Sin. 2012,
36, 270–275. (In Chinese)
[Google Scholar]
15.
Tu ZY. Research of Swimming Performance of Several Typical Fish in the Yalong River. Master’s thesis, Wuhan University, Wuhan, China, 2012. (In Chinese)
16.
Zhou HX, Gan WX. Study on the open feed for
Schizothorax Chongi.
Sci. Aquac. 2016,
11, 67–68. (In Chinese)
[Google Scholar]
17.
Hu XW, Cao WS, Xu SH. The phenomenon of dead eggs in artificial breeding of
Schizothorax chongi and prevention suggestions.
Sci. Aquac. 2017,
39, 5–7. (In Chinese)
[Google Scholar]
18.
Zuo PX, Jin FP, Leng Y, Wang ZF, Wu JX, Deng YL, et al. Prevention and control measures for wheel worm disease in the seedling stage of
Schizothorax chongi.
Aquaculture 2022,
43, 74–75. (In Chinese)
[Google Scholar]
19.
Wang ZF, Deng YL, Xu YR, Zuo PX, Leng Y, Wang C. Acute Toxicity of Ammonia Nitrogen in Different Water Environments to
Schizothorax chongi.
Guizhou Agric. Sci. 2020,
48, 81–84. (In Chinese)
[Google Scholar]
20.
Zhao CY, Wang HL, Zhang DX, Deng YL, Qiu CH, Zheng H, et al. Variation Trend of Fish Community in Upper and Under Longkaikou Dam.
Yunnan Water Power 2022,
38, 224–229. (In Chinese)
[Google Scholar]
21.
Zuo PX, Jin FP, Leng Y, Wang ZF, Wu JX, Yu HM, et al. Comparative analysis of nutritional value evaluation of muscle nutrient composition between wild and cultured
Schizothorax chongi.
Heilongjiang Anim. Sci. Vet. Med. 2023,
18, 126–132. (In Chinese)
[Google Scholar]
22.
Zuo PX, Jin FP, Wang ZF, Wu JX, Zhao JX, Zhao LG, et al. A Preliminary Report on Hybridization and Growth Comparison Experiments of Schizothorax chongi (♀) and Percocypris Pingi Pingi (Tchang) (♂).
Chin. Agric. Sci. Bull. 2024,
40, 159–164. (In Chinese)
[Google Scholar]
23.
Zuo PX, Jin FP, Leng Y, Wang ZF, Wu JX, Yu HM, et al. Anesthetic Effect of Eugenol on Juvenile
Schizothorax chongi.
Chin. Agric. Sci. Bull. 2024,
40, 159–164. (In Chinese)
[Google Scholar]
24.
Guisan A, Tingley R, Baumgartner JB, Naujokaitis‐Lewis I, Sutcliffe PR, Tulloch AI, et al. Predicting species distributions for conservation decisions.
Ecol. Lett. 2013,
16, 1424–1435.
[Google Scholar]
25.
Domisch S, Jaehnig SC, Haase P. Climate‐change winners and losers: Stream macroinvertebrates of a submontane region in Central Europe.
Freshw. Biol. 2011,
56, 2009–2020.
[Google Scholar]
26.
Kuemmerlen M, Schmalz B, Guse B, Cai Q, Fohrer N, Jähnig SC. Integrating catchment properties in small scale species distribution models of stream macroinvertebrates.
Ecol. Model. 2014,
277, 77–86.
[Google Scholar]
27.
Kaelin K, Altermatt F. Landscape-level predictions of diversity in river networks reveal opposing patterns for different groups of macroinvertebrates.
Aquat. Ecol. 2016,
50, 283–295.
[Google Scholar]
28.
Buisson L, Blanc L, Grenouillet G. Modelling stream fish species distribution in a river network: The relative effects of temperature versus physical factors.
Ecol. Freshw. Fish 2008,
17, 244–257.
[Google Scholar]
29.
Pringle C. What is hydrologic connectivity and why is it ecologically important?
Hydrol. Process. 2003,
17, 2685–2689.
[Google Scholar]
30.
Allan JD, Castillo MM, Capps KA. Stream Ecology: Structure and Function of Running Waters; Springer Nature: Cham, Switzerland, 2021.
31.
Nilsson C, Reidy CA, Dynesius M, Revenga C. Fragmentation and flow regulation of the world's large river systems.
Science 2005,
308, 405–408.
[Google Scholar]
32.
Liermann CR, Nilsson C, Robertson J, Ng RY. Implications of dam obstruction for global freshwater fish diversity.
Bioscience 2012,
62, 539–548.
[Google Scholar]
33.
Reid M, Reid M, Thoms M. Ecological significance of hydrological connectivity for wetland plant communities on a dryland floodplain river, MacIntyre River, Australia.
Aquat. Sci. 2016,
78, 139–158.
[Google Scholar]
34.
Lessmann J, Guayasamin JM, Casner KL, Flecker AS, Funk WC, Ghalambor CK, et al. Freshwater vertebrate and invertebrate diversity patterns in an Andean-Amazon basin: implications for conservation efforts.
Neotrop. Biodivers. 2016,
2, 99–114.
[Google Scholar]
35.
Dong X, Li B, He F, Gu Y, Sun M, Zhang H, et al. Flow directionality, mountain barriers and functional traits determine diatom metacommunity structuring of high mountain streams.
Sci Rep. 2016,
6, 24711.
[Google Scholar]
36.
Cote D, Kehler DG, Bourne C, Wiersma YF. A new measure of longitudinal connectivity for stream networks.
Landsc. Ecol. 2009,
24, 101–113.
[Google Scholar]
37.
Bai X, Zhang P, Cao X, Zhang D, Yang Z, Dong X, et al. Incorporating Implicit Information to Disentangle the Impacts of Hydropower Dams and Climate Change on Basin‐Scale Fish Habitat Distribution.
Ecol. Evol. 2024,
14, e70412.
[Google Scholar]
38.
da Mata RA, Tidon R, de Oliveira G, Vilela B, Diniz-Filho JAF, Rangel TF, et al. Stacked species distribution and macroecological models provide incongruent predictions of species richness for Drosophilidae in the Brazilian savanna.
Insect. Conserv. Divers. 2017,
10, 415–424.
[Google Scholar]
39.
Boria RA, Olson LE, Goodman SM, Anderson RP. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models.
Ecol. Model. 2014,
275, 73–77.
[Google Scholar]
40.
Aiello‐Lammens ME, Boria RA, Radosavljevic A, Vilela B, Anderson RP. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models.
Ecography 2015,
38, 541–545.
[Google Scholar]
41.
Steen VA, Tingley MW, Paton PW, Elphick CS. Spatial thinning and class balancing: Key choices lead to variation in the performance of species distribution models with citizen science data.
Methods Ecol. Evol. 2021,
12, 216–226.
[Google Scholar]
42.
Zhang P, Dong X, Grenouillet G, Lek S, Zheng Y, Chang J. Species range shifts in response to climate change and human pressure for the world's largest amphibian.
Sci. Total Environ. 2020,
735, 139543.
[Google Scholar]
43.
Hijmans RJ, Cameron SE, Parra JL, Jones PG, Jarvis A. Very high resolution interpolated climate surfaces for global land areas.
Int. J. Climatol. 2005,
25, 1965–1978.
[Google Scholar]
44.
Harris I, Osborn TJ, Jones P, Lister D. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset.
Sci. Data. 2020,
7, 109.
[Google Scholar]
45.
Fick SE, Hijmans RJ. WorldClim 2: New 1‐km spatial resolution climate surfaces for global land areas.
Int. J. Climatol. 2017,
37, 4302–4315.
[Google Scholar]
46.
De Marco PJ, Nóbrega CC. Evaluating collinearity effects on species distribution models: An approach based on virtual species simulation.
PLoS ONE 2018,
13, e0202403.
[Google Scholar]
47.
Grenouillet G, Buisson L, Casajus N, Lek S. Ensemble modelling of species distribution: the effects of geographical and environmental ranges.
Ecography 2011,
34, 9–17.
[Google Scholar]
48.
Rosenblatt F. The perceptron: a probabilistic model for information storage and organization in the brain.
Psychol. Rev. 1958,
65, 386–408.
[Google Scholar]
49.
Booth TH, Nix HA, Busby JR, Hutchinson MF. BIOCLIM: The first species distribution modelling package, its early applications and relevance to most current MAXENT studies.
Divers. Distrib. 2014,
20, 1–9.
[Google Scholar]
50.
Hastie T, Tibshirani R, Buja A. Flexible discriminant analysis by optimal scoring.
J. Am. Stat. Assoc. 1994,
89, 1255–1270.
[Google Scholar]
51.
Phillips SJ, Anderson RP, Schapire RE. Maximum entropy modeling of species geographic distributions.
Ecol. Model. 2006,
190, 231–259.
[Google Scholar]
52.
Thuiller W, Lafourcade B, Engler R, Araújo MB. BIOMOD–A platform for ensemble forecasting of species distributions.
Ecography 2009,
32, 369–373.
[Google Scholar]
53.
Swets JA. Measuring the accuracy of diagnostic systems.
Science 1988,
240, 1285–1293.
[Google Scholar]
54.
Allouche O, Tsoar A, Kadmon R. Assessing the accuracy of species distribution models: prevalence, kappa and the true skill statistic (TSS).
J. Appl. Ecol. 2006,
43, 1223–1232.
[Google Scholar]
55.
Franklin J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2009.
56.
Yang M, Zhao H, Xian X, Liu H, Li J, Chen L, et al. Potential global geographical distribution of
Lolium temulentum L. under climate change.
Front. Plant Sci. 2022,
13, 1024635.
[Google Scholar]
57.
Araújo MB, New M. Ensemble forecasting of species distributions.
Trends Ecol. Evol. 2007,
22, 42–47.
[Google Scholar]
58.
Buisson L, Thuiller W, Casajus N, Lek S, Grenouillet G. Uncertainty in ensemble forecasting of species distribution.
Glob. Chang. Biol. 2010,
16, 1145–1157.
[Google Scholar]
59.
Warren ML, Jr., Pardew MG. Road crossings as barriers to small-stream fish movement.
Trans. Am. Fish. Soc. 1998, 127, 637–644.
[Google Scholar]
60.
Oldford G, Cote D, Kehler DG, Riefesel GR, Wiersma YF. FIPEX v10. 4: An ArcGIS Desktop Add-in for assessing impacts of fish passage barriers and longitudinal connectivity of rivers.
SoftwareX 2023, 23, 101469.
[Google Scholar]
61.
Caissie D. The thermal regime of rivers: a review.
Freshw. Biol. 2006,
51, 1389–1406.
[Google Scholar]
62.
Dala‐Corte RB, Giam X, Olden JD, Becker FG, Guimarães TdF, Melo AS. Revealing the pathways by which agricultural land‐use affects stream fish communities in South Brazilian grasslands.
Freshw. Biol. 2016,
61, 1921–1934.
[Google Scholar]
63.
Keppeler FW, de Souza AC, Hallwass G, Begossi A, de Almeida MC, Isaac VJ, et al. Ecological influences of human population size and distance to urban centres on fish communities in tropical lakes.
Aquatic. Conserv. Mar. Freshw. Ecosyst. 2018,
28, 1030–1043.
[Google Scholar]
64.
Su G, Logez M, Xu J, Tao S, Villéger S, Brosse S. Human impacts on global freshwater fish biodiversity.
Science 2021, 371, 835–838.
[Google Scholar]
65.
Wu J, Xu HG, Chen L. A Review of Impacts of Climate Change on Species.
J. Ecol. Rural. Environ. 2011, 27, 1–6. (In Chinese)
[Google Scholar]
66.
Sun HY, Xue LF, Jiang H, Chen YF. Fish-Based Evaluation of Protected Areas in the Jinsha River Basin.
Environ. Impact Assess. 2019,
41, 13–16. (In Chinese)
[Google Scholar]
67.
Hermoso V, Filipe AF, Segurado P, Beja P. Freshwater conservation in a fragmented world: Dealing with barriers in a systematic planning framework.
Aquatic. Conserv. Mar. Freshw. Ecosyst. 2018,
28, 17–25.
[Google Scholar]
68.
Hermoso V, Kennard MJ, Linke S. Integrating multidirectional connectivity requirements in systematic conservation planning for freshwater systems.
Divers. Distrib. 2012,
18, 448–458.
[Google Scholar]
69.
Yujun Y, Yanning G, Shanghong Z. The impact of dams on the river connectivity of the two largest river basins in China.
River Res. Appl. 2022,
38, 185–193.
[Google Scholar]
70.
Chu C, Mandrak NE, Minns CK. Potential impacts of climate change on the distributions of several common and rare freshwater fishes in Canada.
Divers. Distrib. 2005, 11, 299–310.
[Google Scholar]
71.
Ramankutty N, Evan AT, Monfreda C, Foley JA. Global Agricultural Lands: Croplands, 2000; NASA Socioeconomic Data and Applications Center (SEDAC) [Dataset]: Palisades, NY, USA, 2010.
72.
Ramankutty N, Evan AT, Monfreda C, Foley JA. Farming the Planet: 1. Geographic Distribution of Global Agricultural Lands in the Year 2000.
Glob. Biogeochem. Cycles. 2008,
22, GB1003.
[Google Scholar]
73.
Oakleaf JR, Kennedy CM, Baruch-Mordo S, West PC, Gerber JS, Jarvis L, et al. Development Threat Index; NASA Socioeconomic Data and Applications Center (SEDAC) [Dataset]: Palisades, NY, USA, 2019.
74.
Oakleaf JR, Kennedy CM, Baruch-Mordo S, West PC, Gerber JS, Jarvis LJ, et al. A World at Risk: Aggregating Development Trends to Forecast Global Habitat Conversion.
PLoS ONE 2015,
10, e0138334.
[Google Scholar]
75.
Wang P, Huang C, Brown de Colstoun EC, Tilton JC, Tan B. Global Human Built-up And Settlement Extent (HBASE) Dataset From Landsat; NASA Socioeconomic Data and Applications Center (SEDAC) [Dataset]: Palisades, NY, USA, 2017.
76.
Center for International Earth Science Information Network—CIESIN—Columbia University. Gridded Population of the World, Version 4 (GPWv4): Population Count, Revision 11; NASA Socioeconomic Data and Applications Center (SEDAC) [Dataset]: Palisades, NY, USA, 2018.
77.
Center for International Earth Science Information Network—CIESIN—Columbia University, Centro Internacional de Agricultura Tropical—CIAT. Gridded Population of the World, Version 3 (GPWv3): Population Density Grid; NASA Socioeconomic Data and Applications Center (SEDAC)[Dataset]: Palisades, NY, USA, 2005.
78.
Wildlife Conservation Society—WCS, Center for International Earth Science Information Network—CIESIN—Columbia University. Last of the Wild Project, Version 2, 2005 (LWP-2): Global Human Influence Index (HII) Dataset (Geographic); NASA Socioeconomic Data and Applications Center (SEDAC) [Dataset]: Palisades, NY, USA, 2005.